Адрес этой статьи в интернете: www.biophys.ru/archive/congress2012/proc-p21-d.htm
Влияние вариаций изотопного состава воды на ее биологическую активность.
Лобышев В.И., Киркина А.А.
Физический факультет, Московский государственный университет им. М.В.Ломоносова,
119991, Россия, Москва, Ленинские горы. E-mail: lobyshev@yandex.ru
После первых работ Льюиса и Тейлора [1-2] с концентрированной тяжелой водой D2O стало ясно, что она ингибирует жизненно важные процессы. Это было подтверждено другими исследователями и сформировалось устойчивое мнение, что тяжелая вода несовместима с жизнью. Так началось новое направление – исследование изотопных эффектов тяжелой воды в биологических системах, которое не исчерпано и по настоящее время. Эволюцию взглядов в этом направлении с анализом физико-химических механизмов изотопных эффектов можно найти в [3]. В целом, первоначальная точка зрения оказалась верной и биологические изотопные эффекты тяжелой воды высокой концентрации можно назвать ингибирующими. Наряду с влиянием концентрированной тяжелой воды, несомненный интерес представляет изучение влияния вариации малых концентраций дейтерия на живые организмы. Первые работы в этой области были выполнены еще в 30-е годы, но они долгое время не возобновлялись и не цитировались. Экспериментально было показано, что небольшие изменения естественного изотопного состава воды, могут приводить к неожиданно большим эффектам, часто противоположным действию концентрированной тяжелой воды.
Напомним, что природная вода является смесью изотопных форм молекул вследствие наличия стабильных изотопов 16О, 17О, 18О, 1Н, 2Н и радиоактивного изотопа 3Н с периодом полураспада 12,5 лет. Радиоактивные изотопы 14О, 15О, 19О характеризуются временами полураспада 76,5; 118 и 29,5 с и практически отсутствуют в природной воде. Относительное содержание наиболее распространенных изотопов дейтерия и 18О в природных водах составляет в среднем 0,015% (150 ppm) и 0,2% и варьирует в пределах 0,0079 – 0,0195% и 0,1887 – 0,2083% соответственно, вследствие фракционирования изотопов воды при фазовых переходах пар-жидкость-лед (снег), при сорбции и фильтрации в природных процессах [4].
Эксперименты с водой, слегка обогащенной дейтерием, были начаты Банесом в 1933 г. [5-12]. Исследовано действие воды, полученной электролизным методом, с содержанием дейтерия 0,06% на развитие водоросли Spirogyra nitida, культивируемой под рассеянным светом при температуре 10-140С. Для филаментов водоросли в воде с повышенным содержанием дейтерия (в дальнейшем для краткости будем использовать термин «утяжеленная» вода) было характерно уменьшение подвижности, значительное уменьшение количества разрывов и увеличение длительности жизни [6]. В контрольном опыте в дистиллированной воде филаменты рвались на более мелкие фрагменты, а их распределение по длинам было более широким. В одном из опытов филаменты в «утяжеленной» воде вовсе не имели разрывов в течение полутора месяцев [12]. Разрывы наблюдали лишь в длинных филаментах, но не в коротких – использованных для дальнейших количественных исследований.
В обычной воде у 322 клеток, содержащихся в 15 коротких филаментов из 5-50 клеток, средняя продолжительность жизни одной клетки составила 3 дня, в то время как в «утяжеленной» воде у 355 клеток, содержащихся в 16 филаментах, средняя продолжительность жизни клеток составляла 6 дней. Увеличенная продолжительность жизни в «утяжеленной» воде наблюдалась во всех случаях, независимо от средней длины клетки. В то же время, увеличение концентрации дейтерия в воде не приводило к заметным изменениям в скорости деления клеток или их удлинения, однако вследствие большей долговечности клеток наблюдалось большее число делений клеток [10]. Особо хочется подчеркнуть, что многократная (до 6 раз) перегонка воды не изменила получаемых результатов. Это дает основание утверждать, что наблюдаемые эффекты не обусловлены наличием посторонних примесей в «утяжеленной» воде, появившихся в процессе электролиза.
В работе [9] было установлено, что «утяжеленная» вода с малой концентрацией дейтерия оказывает влияние на продолжительность жизни Spyrogira только в фотосинтезирующих водорослях на свету, но не в темноте. Из 1088 клеток водоросли в 0,47% D2O через два дня при освещенности фут-свечей (примерно 646 лк) развивается 72% клеток, тогда как только 18% из 1129 клеток оставались живыми в темноте. В обычной воде 32% из 1266 развивались на свету той же интенсивности и 16% из 789 клеток в темноте. На основании этого эксперимента авторы делают вывод о том, что «утяжеленная» вода способствует процессу фотосинтеза.
В экспериментах, проведенных Фоксом и др.[18], с морскими диатомовыми водорослями Nitzchia bilobata в морской воде, содержащей 1% дейтерия, было обнаружено 16-ти процентное увеличение лаг - периода на кривых роста. Для другой водоросли N. closterium отмечается противоположная тенденция, однако, этот результат статистически не достоверен. Изменений стационарной концентрации диатомовых в воде с различным изотопным составом не обнаружено.
Наряду с ингибирующим (или стабилизирующим) влиянием «утяжеленной» воды было обнаружено также стимулирующее действие на живые организмы. Так количество клеток Euglena gracilus, культивируемых в течение 45 суток при температуре 17-200С, было увеличено на 15,6% в среде, содержащей 0,06% дейтерия. При этом количество подвижных микроорганизмов по сравнению с контролем возрастало примерно в 2,3 раза [7].
Аномальное влияние «утяжеленной» воды было подтверждено Ричардсом в опытах с низшими грибами [13-14]. Исследуя Saccharomyces cerevisae в среде Вильямса в электролизной воде с содержанием 0,06% дейтерия за 143 часа опыта, он не обнаружил различий между контролем и опытом в числе клеток на единицу объема, в количестве почек, числе мертвых или поврежденных клеток, проницаемых для метиленового синего. В то же время в «утяжеленной» воде общий объем клеток после центрифугирования в мерных пробирках был на 20% больше, чем в контроле, хотя средний размер клеток в опыте лишь на 3% больше. Клетки в «утяжеленной» воде были более однородны по размеру – коэффициент вариации формы на 8% меньше, чем в контроле. Сухой вес дрожжей оказался больше на 26%, что свидетельствует о большем количестве синтезированного твердого вещества в клетках, выросших в среде с повышенным содержанием дейтерия. В работе [14], наряду с вышеуказанными результатами, отмечается, что дрожжи, выращенные в «утяжеленной» воде, содержат больше гликогена, но меньше азота. Наибольшее влияние малых добавок дейтерия в среде наблюдали во время быстрой клеточной пролиферации, когда соотношение средней поверхности клеток к их объему максимально.
Еще более поразительные результаты были получены Майером [15, 16]. После выращивания в течение 5 дней чистой культуры Aspergillus sp. в инкубаторе при температуре 370С в среде Пфеффера сухой вес мицелия, выросшего в среде, содержащей 0,47% дейтерия, оказался в 16 раз больше, чем в контроле. Отмечены также морфологические изменения культур, выросших в средах с различным изотопным составом. В обычных условиях плесень имеет плоскую форму и равномерно распределенные плодовые тела. В «утяжеленной» воде плесень сильно завита, имеет провалы и напоминает рельеф коры головного мозга, плодовых тел существенно меньше, а их распределение по поверхности очень неравномерно.
Исследование выделения газа при росте пекарских дрожжей в растворе Пастера, содержащем 0,25 и 0,5% D2O, не дали столь больших эффектов. В конце ферментации количество газа в 0,5% D2O было уменьшено на 6-6,5% [20].
Банес и др. [10] не нашли изменений в регенерации плоских червей Phagocata gracilis в 0,06%-ной тяжелой воде. Однако через несколько месяцев можно было наблюдать качественное различие. В обычной воде планарии сморщились примерно до 1/5 от исходной длины тела, тогда как в «утяжеленной» воде сокращение было незначительным. Опыты с планариями были повторены с большей концентрацией дейтерия – 0,47% и был обнаружен новый эффект. На планариях в этой воде быстро паразитировал плесневый грибок, и животные погибли в течение трех недель [21]. В некоторых опытах живые животные покрывались слизью, либо пучками мицелия. В этой же работе аналогичное ускорение роста плесневых грибов обнаружено в экспериментах с прорастающими семенами Aquilegia. На семенах, замоченных «утяжеленной» водой появилась большая масса мицелия белого цвета. Этот гриб в основном сапрофитен, т.к. появился, в основном, на не проросших семенах. Судя по приведенным фотографиям, контрольные животные и семена были свободны от паразитов. Результаты этой работы хорошо согласуются с данными Майера об активации роста плесени в «утяжеленной» воде.
Пытаясь понять механизм действия малых концентраций дейтерия на живые организмы, Банес провел ряд экспериментов на молекулярном уровне [12]. Ему удалось обнаружить замедление гидролиза крахмала панкреатической амилазой и ингибирование зимазного комплекса в воде, содержащей 0,06% дейтерия. После 24-часовой инкубации амилазы в «утяжеленной» воде стадия эритродекстрина достигалась за 8 минут, а в контроле за 6 минут. Преинкубация зимазного комплекса в «утяжеленной» воде приводила к 10%-ному уменьшению количества СО2, причем этот эффект не менялся при увеличении времени преинкубации фермента от 16 до 166 часов. Отмечалось также, что влияние дейтерия не проявляется при проведении реакции без преинкубации фермента или при инкубации одного лишь субстрата. Банес считал, что обнаруженные изменения ферментативной активности могут быть возможной причиной влияния малых концентраций дейтерия на живые организмы. Описывая работы Банеса, следует сказать также о том, что он с 1932 г. активно исследовал влияние талой и конденсированной воды из пара на живые организмы. Основной причиной, объясняющей различное биологическое действие этих вод, он считал сохранение структурированности в талой воде и ее полное разрушение в конденсированной воде.
Влияние 0,11% тяжелой воды испытано на мышах, зараженных саркомой [19]. Из 100 мышей с инокулированной опухолью 50 штук в течение 10 дней получали инъекции по 1см3 «утяжеленной» воды ежедневно внутрь опухоли и вокруг нее. Контрольная группа мышей получала инъекции обычной воды. В большинстве случаев различий в развитии опухолей не обнаружено, однако, в 12% случаев наблюдали ускоренный рост и увеличение размеров опухоли в опытной серии. Авторы работы полагали, что этот факт нельзя объяснить случайными причинами и связывали его с активирующим влиянием «утяжеленной» воды.
Наряду с перечисленными, существуют такие работы, в которых влияние малых добавок тяжелой воды не обнаружено. Так, Махт и Дэвис [17] в воде с концентрацией дейтерия 0,05% наблюдали лишь небольшое замедление прорастания семян и роста проростков люпина. Они не обнаружили отличий от контроля в ряде биологических процессов, а именно: в ферментации пекарских дрожжей; в реакции мышей на инъекции физиологического раствора; в поведении золотой рыбки; в ритмических сокращениях гладкой мускулатуры полоски кишечника кошки; в индуцированных различными фармакологическими препаратами сокращениях образцов гладких мышц из кишечника кошки, стенок сосудов белых крыс, стенок матки морской свинки; в давлении крови и дыхании наркотизированных кошек при внутривенных инъекциях физиологического раствора, а также при действии тинктуры наперстянки. К сожалению, сравнить эти результаты с данными других авторов, например Ричардса, невозможно, поскольку в этой работе полностью отсутствует описание постановки эксперимента.
Добавленная D2O в количестве 2,5.10–4% не изменила характеристик роста популяции водяных блох [22]. Не наблюдалась стимуляции прорастания семян гороха Pisum sativum [23]. В 0,46-процентной тяжелой воде не было замечено количественных или качественных различий в следующих процессах: прорастании милдью пшеницы Erysiphe graminis tritici, скорости роста корешков проростков пшеницы, скорости дыхания проростков пшеницы, а также, в противоположность Майеру, в росте плесени Aspergillus niger [24]. «Утяжеленная» вода в концентрации 0,13; 0,7 и 5% не изменила морфологии и скорости размножения молочнокислых бактерий Lactobacillus acidophilus и L. Bulgaricus [25]. Незначимый результат получен также Фоксом [26] при исследовании влияния 1-процентной тяжелой воды на активность каталазы и амилазы мышц и скорость гидролиза глюкозида амигдалина эмульсией из горького миндаля. Изменений в ферментативной активности не было обнаружено как сразу после добавления тяжелой воды, так и при предварительной инкубации фермента и субстрата в растворе «утяжеленной» воды в течение нескольких часов.
Чаще всего авторы работ, получивших незначимые результаты, видят причину появления эффектов малых концентраций дейтерия в других работах за счет посторонних примесей. Это мнение, хотя и может иметь отношение к некоторым работам, следует признать ошибочным. Достаточно вспомнить уже цитированную работу [6], где было показано, что многократная перегонка тяжелой воды не привела к изменению результатов. Кроме того, в ответ на замечания оппонентов, в специально выполненных работах было показано, что стимуляция роста плесени, растущей в воде, обогащенной дейтерием, не может быть обусловлена, например, примесями парафина [8, 15].
Наиболее тщательно, на наш взгляд, выполнена работа [27], в которой содержится также анализ способа приготовления воды. Показано, что для исключения дополнительных ошибок и обеспечения одинакового рН необходимо дистиллировать обычную и «утяжеленную» воду и стерилизовать растворы в одном, желательно стеклянном приборе. В противном случае растворы имеют различающиеся значения рН, что приводит к существенным изменениям результата в контрольных экспериментах. Были исследованы водоросли – Lemna minor, Chlamidomonas, Phormidium autumnale; безхлорофилльные организмы: грибы –Rhizopus nigricans, Penicillium eitrinum; бактерии – B. cloacae, B.subtilis, Sarcina lutea; дрожжи – B. cerevisiae. Показано, что в воде, содержащей 0,1% дейтерия, значительно активируется развитие водорослей (у Lemna активация выражена слабо), сильно угнетается рост как скороспелых, так и не скороспелых бактерий, несколько ускоряется рост дрожжей. Вода, содержащая 0,13% дейтерия, значительно замедляет развитие проростков овса, но их реакция на освещение такая же, как в контроле. В этой же работе было проверено влияние разбавленной тяжелой воды на подвижность микроскопических водяных змей Ancylus tasmanicus длиной 2-3 мм. В воде, содержащей 0,13% дейтерия, подвижность змей уменьшалась через 2-3 дня и через 3 недели движения практически полностью прекращались. Этот феномен оказался обратимым. После смены «утяжеленной» воды на обычную, через 3 дня змеи ползали и активно питались.
На этом литературные источники 30-х годов, описывающие влияние малых добавок тяжелой воды на живые организмы, исчерпываются. После длительного перерыва аналогичные работы были возобновлены в 60-х годах. В работах Б.Н.Родимова с сотр. [28-30] было показано активирующее влияние талой воды, полученной из снега, на ряд биологических процессов и было получено увеличение продуктивности сельскохозяйственных животных и растений. В эксперименте с двумя группами по 14 кур несушек, пивших обычную и снеговую воду, было получено, что за время с декабря по март среднее количество яиц на одну несушку и общий вес яиц в опытной группе в 2 раза больше, чем в контрольной. Отмечается, что хотя подопытные куры пили талой воды больше, корма они потребляли столько же, сколько контрольные. За это же время суммарный привес поросят в опытной группе оказался в 1,4 раза больше, чем в контрольной. Сила роста семян пшеницы, замоченных талой водой была на 41% больше, чем в контроле. Огурцы, которые поливали талой водой после высадки рассады дали урожай 210%, а те у которых замачивали еще и семена – 290%, урожай редиса составил 230% [28].
К этому времени уже существовала хорошо разработанная теория фракционирования изотопов при фазовых переходах воды. Накопилось также много экспериментальных данных, свидетельствующих о том, что атмосферные осадки содержат уменьшенной количество тяжелых изотопов воды и в первую очередь дейтерия. Наиболее «облегченной» является вода из свежевыпавшего снега. Поэтому рабочей гипотезой, объясняющей влияние талой воды на животных и растения, стало стимулирующее влияние пониженной концентрации дейтерия в воде. Однако, оставалась еще альтернативная гипотеза об особых свойствах “структурированной” талой воды, существующая еще с 30-х годов. Последующий эксперимент позволил устранить эту альтернативу. Три группы семян ячменя замачивали около 8 часов в разных водах – талой снеговой, талой с добавлением недостающего количества тяжелой воды (0,003%) и контрольной водопроводной [29]. Затем семена подсушивали и высевали. Второй и третий варианты опыта дали сходные результаты. В первом опыте со снеговой водой вес зерна оказался на 18%, а биологическая урожайность на 25% больше. Аналогичный полевой эксперимент с пшеницей дал еще более убедительное подтверждение (22 и 56% соответственно) гипотезы о стимулирующем влиянии воды с пониженной концентрацией дейтерия. Эти эксперименты показали также, что возможное объяснение наблюдаемых эффектов за счет не идентифицированных высокоактивных примесей несостоятельно, поскольку стимулирующий эффект снимался добавлением небольшого количества тяжелой воды.
В работе [30] представлены результаты исследований разнообразных биологических объектов, где было показано, что снеговая вода задерживает старение культур Хела, Хеп-2, детройт-6, 580, печени и фибробластов куриных эмбрионов. Даже через 7-8 суток не было замечено деградации и еще наблюдали рост культур. Был проведен также опыт с повышенным до 3% содержанием дейтерия в питьевой воде мышей. Контролем в этом случае была обычная вода. Вес новорожденных мышат в этом опыте был на 20% меньше, а среди мышат преобладали самцы (около 75%). Мышата росли медленнее, половое развитие у них запаздывало. Самки второго поколения оказались неспособными к лактации и их мышата погибали. Третьего поколения получить не удалось. Было показано также, что дейтерий аккумулируется в яичниках белых мышей, что приводит к морфологическим изменениям на всех стадиях развития яйцеклеток. Использование талой воды в качестве питьевой привело к качественно противоположным результатам, а развитие яйцеклеток стимулируется водой с пониженной концентрацией дейтерия.
Качественно похожие результаты были получены в опытах с мухами дрозофилами. В среде, содержавшей водопроводную воду, количество самцов и самок с точностью 0,5% было одинаковым. В среде с «утяжеленной» водой, содержавшей 2 и 3% дейтерия, соотношение самцов и самок оказалось равным 56:44 и 53:47 соответственно. Напротив, в среде, содержавшей снеговую воду и воду с искусственно пониженной концентрацией дейтерия, количество самок возрастало так, что соотношение самцов и самок стало 47:53. Погрешность эксперимента не приводится, указано только, что в одной серии экспериментов было проанализировано 1,5 тысячи особей. Еще более убедительные результаты были получены при анализе влияния снеговой воды на соотношение полов в трех поколениях. Результаты эксперимента, выполненного на 32 тысячах особей, представлены в таблице 1.
|
Таблица 1 |
||||
|
Поколения |
1 |
2 |
3 |
Контроль |
|
Самцы, % |
41,4 |
40,7 |
39,6 |
48,0 |
|
Самки, % |
58,6 |
59,3 |
60,4 |
52,0 |
Видно, что уменьшение концентрации дейтерия ведет к преобладанию в потомстве самок, причем этот эффект усиливается в последующих поколениях.
Аналогично описанному выше эксперименту с ячменем и пшеницей были поставлены опыты с хлореллой [30]. После культивирования получили среднее увеличение массы хлореллы в снеговой воде на 88% и уменьшение на 27% в снеговой воде с добавлением 0,003% D2O относительно массы хлореллы в контрольном опыте. Таким образом, еще раз подтверждается именно специфичность действия малых концентраций дейтерия в воде.
Авторы работы [30] обратили также внимание на возможные демографические последствия вариаций изотопного состава питьевой воды. Так, например, число долгожителей с возрастом более 100 лет на 106 человек в среднем по РФ составляет 81 человек, в то время, как в Чечено-Ингушетии – 353, а в Якутии 324 человека. Первые из группы долгожителей пьют талую воду из горных рек, а вторые снеговую воду длительное время в течение года. Конечно, приведенные статистические данные можно объяснять и другими причинами, но отмеченная корреляция, безусловно, заслуживает внимания.
Известна еще одна работа [31], в которой показано, что овес быстрее прорастает в воде, полученной из антарктического льда, облегченного по кислороду на 49% и по дейтерию на 400 % относительно стандарта SMOW.
Результаты цитированных работ убедительно показывают, что тяжелая вода в малых количествах может оказывать различное влияние, зависящее как от вида организма, так и от исследуемой функции. При этом может наблюдаться как активирование, так и ингибирование реакций исследуемой системы. Эти наблюдения убедительно подтверждают широкую вариабельность ответов различных организмов на небольшое изменение изотопного состава воды и снимают кажущуюся противоречивость результатов, полученных разными авторами. Феномен аномального влияния малых вариаций концентрации дейтерия в воде на развитие ряда живых организмов был установлен еще в начале тридцатых годов, однако он был не понят и незаслуженно забыт.
На наш взгляд, основным недостатком вышеописанных работ является отсутствие зависимостей наблюдаемых эффектов от концентрации дейтерия в воде. Очевидно, что эта зависимость должна быть немонотонна, поскольку большие концентрации дейтерия всегда приводят к ингибированию, а малые могут оказывать активирующее действие. Поэтому выбор одной произвольной концентрации дейтерия в опытах с организмами, имеющими различную чувствительность к изменениям изотопного состава воды, может привести к различным, в том числе и незначимым результатам.
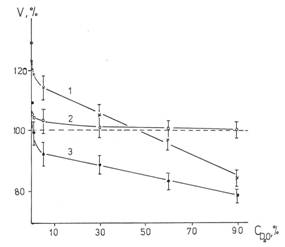
Наши исследования гидролитической активности Na,K-AТФазы из мозга быка, проведенные в широком интервале значений концентраций добавленной D2O обнаружили аномальное активирующее влияние малых добавок D2O к исследуемой системе при 370С. Было показано [32, 33], что активность фермента линейно зависит от концентрации дейтерия в области 5-90%, но с уменьшением концентрации изотопный эффект меняет знак и резко возрастает (рис.1). Кривые нормированы на значение активности АТФазы в Н2О. Заметим, что величина изотопного эффекта зависит и от соотношения ионов натрия и калия в среде при постоянной ионной силе среды (рис.2). В области относительно малых концентраций натрия (10-60 мМ) и малых концентраций калия (10-50 мМ) изотопный эффект максимален, а в области промежуточных концентраций ионов в воде, содержащей более 30% дейтерия, он практически отсутствует. Из рис.2 видно также, что характер изотопных эффектов больших и малых концентраций дейтерия качественно различается. Переход к полулогарифмической зависимости (рис.3) свидетельствует, что при малых, но увеличенных относительно природного содержания,
|
|
|
|
|
Рис. 1. Зависимость скорости гидролиза АТФ от концентрации дейтерия в воде при 1-50мМ NaCl / 100мМ KCl, 2- 70/80 3- 135/15. T=370C. pHapp=7,4.
|
Рис. 2. Зависимость гидролитической активности от концентрации дейтерия 1- H2O, 2- 0,05% D2O, 3- 30%, 4- 60%, 5- 90%. pHapp=7,4. T= 370С. |
|
|
|
|
|
|
Рис. 3. Зависимость скорости гидролиза АТФ от концентрации дейтерия в воде при 70мМ NaCl, 80мМ KCl, T=370C pHapp=7,4 |
Рис. 4. Зависимость гидролитической активности от концентрации дейтерия 1- H2O, 2- 0,05% D2O, 3- 60%, 4- 90%, T=130C |
|
концентрациях дейтерия в воде ингибирование фермента сменяется его сильной активацией. Можно было бы думать, что эффект малых концентраций обусловлен наличием примесей, однако этот эффект сохраняется после дополнительной очистки D2O перегонкой в вакууме. Более того, он исчезает при уменьшении температуры ниже температуры фазового перехода липидов в липидно-белковом комплексе, равной примерно 200С (рис. 4). При этом величина изотопных эффектов больших концентраций D2O возрастает в соответствие с теорией. При замене ионов калия в среде на ионы цезия при температуре 370С наблюдается ингибирование ферменты высокими концентрациями D2O и активация малыми, как и в среде с калием. Активация гидролитической активности малыми добавками D2O была обнаружена нами также на мембранном препарате Сa,Mg–АТФазы, однако в препарате миозина из мышц кролика в интервале 0,02-1% D2O аномальный активирующий эффект не наблюдался. Ранее в работе [34] на основании анализа изотопных эффектов D2O высокой концентрации в миозине и Na,K–АТФазе был сделан вывод об идентичности механизма гидролиза АТФ этими белками. Результаты, полученные в области малых концентраций дейтерия, не согласуются с этой точкой зрения. Наблюдаемая активация связана не с локальным химическим механизмом реакции, а с коллективными свойствами комплекса липид-белок-вода. Удовлетворительного объяснения этого феномена до сих пор нет. Хотя известны факты фракционирования изотопов воды в гидратных оболочках ионов и биополимеров, а также некоторых сорбентов, константы фракционирования невелики, да и теоретический предел для ОН-группы близок к 10. Поэтому предложить высокую специфичность локальных групп к следовым концентрациям дейтерия не представляется возможным. Отметим только, что отношение числа атомов дейтерия к числу молекул Na,K–АТФазы в области максимальной активации примерно равно 103, а отношение числа атомов дейтерия к числу атомов протия в природной воде примерно 10–4.
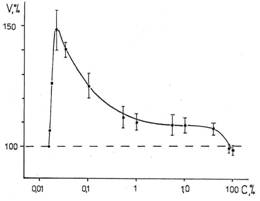
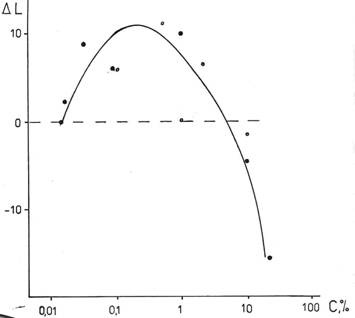
Эксперименты по регенерации гидроидных полипов Obelia geniculata, проведенные в широком интервале значений концентраций добавленной D2O к обычной морской воде показали, что наряду с резким ингибированием процесса высокими концентрациями дейтерия, наблюдается активация регенерации малыми концентрациями, что иллюстрируется рис.5 [35]. Точки разной жирности получены в разные годы.
Вследствие значительной вариабельности результатов, трудно указать точное значение концентрации дейтерия, вызывающей максимальный активирующий эффект. Можно лишь ориентировочно сказать, что она близка к 0,1%, что примерно в 6 раз превосходит среднюю величину концентрации дейтерия в морской воде.
|
|
Рис. 5. Влияние различной концентрации дейтерия в воде на длину регенерировавшей части гидроида к 72-му часу в относительных единицах. ΔL = LD – LH
|
Концентрация дейтерия, соответствующая максимальной активации процесса регенерации и Na,K–АТФазы лишь на порядок превосходит величину концентрации дейтерия в природной воде. Учитывая близость этих величин можно высказать предположение о биологической значимости вариаций изотопного состава воды, сопровождающих фракционирование воды в природе. Эта мысль не нова и обсуждалась еще Банесом, а затем Родимовым в связи с проблемой активации биосистем талой водой. В этих работах, однако, использовалась лишь одна концентрационная точка. Поскольку в процессе генезиса морского дрейфующего льда наблюдаются изменения его изотопного состава, это дает уникальную возможность исследования возможных связей между изотопным составом льда и развитием одноклеточных водорослей.
|
|
|
|
Рис. 6. Вариации изотопного состава кислорода в толще арктического дрейфующего льда СП–23. Координата “0” соответствует верхней кромке льда. |
Рис. 7. Корреляция относительной концентрации каротиноидов (в отн. ед) с изотопным составом кислорода в образцах льда.
|
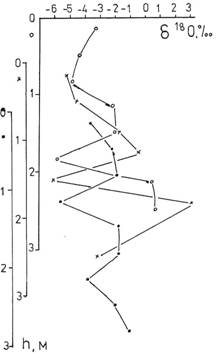
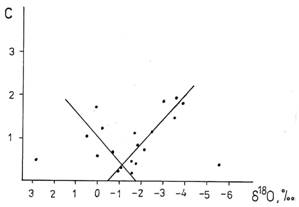
Эксперименты были проведены на дрейфующей полярной станции СП–23 [36, 37]. Колонки льда диаметром 18см и длиной, равной толщине многолетнего льда (около 3м), вырезали на небольшой площади для уменьшения систематической ошибки, обусловленной различиями в структуре льда. Вмораживание мерной рейки в лед после взятия первой пробы обеспечило возможность определения абсолютного значения координаты высоты льда. Из колонки льда получали образцы льда длиной около 30 см. В этих образцах измеряли соленость, концентрации фосфата, кремния, хлорофилла, каротиноидов, содержание тяжелого изотопа кислорода 18О. Изотопные измерения проводили методом масс – спектрометрии [38]. Содержание 18О выражали в единицах промилле от международного стандарта SMOW по формуле δ18О = (R/RSMOW – 1)· 103, ‰ где R=[18O]/[16O]. По техническим причинам мы не могли измерять концентрацию дейтерия в воде, но измеряли концентрацию 18О и использовали линейную корреляцию δD = 8δ18О − 10 (линию Крейга) между концентрацией тяжелых изотопов водорода и кислорода [4]. Результаты изотопного анализа представлены на рис. 6. Координата верхней точки смещается вследствие летнего таяния льда, а приращение снизу происходит из-за намерзания льда снизу. Уменьшение содержания 18О объясняется намерзанием талой снеговой воды, проникающей под лед, а увеличение – намерзанием льда из морской воды. Так реализуется природный пространственно-временной периодический “фракционный генератор “, обеспечивающий вариации изотопного состава в толще льда. Во льду развиваются сине-зеленые и зеленые одноклеточные водоросли. Количество наиболее древних сине-зеленых водорослей определяли по концентрации каротиноидов, а зеленых по концентрации хлорофилла. Была проанализирована возможная связь изотопного состава с концентрацией каротиноидов. Результаты представлены на рис. 7. Точки, соответствующие 2,8 и -5,6% исключены из рассмотрения. Оставшийся массив точек описывается двумя линиями регрессии с коэффициентами корреляции, отличающимися от нуля при 1-процентном уровне значимости (левая ветвь) и даже 0,1-процентном уровне значимости (правая ветвь). Обращает на себя внимание точка пересечения линий регрессии. Она соответствует δ18О =–1,2‰ и весьма близка к содержанию в воде океана третичного периода δ18О =–1,3‰ [39]. В этой точке концентрация каротиноидов, отражающая количество сине-зеленых водорослей минимальна. Относительно этой точки, как уменьшение, так и увеличение концентрации 18О, а значит и дейтерия, коррелирует с ростом концентрации сине-зеленых водорослей. Хотя для хлорофилла такой четкой картины не было, анализ распределения средних концентраций хлорофилла и каротиноидов однотипны и характеризуются двумя максимумами. “Облегчение” и “утяжеление” изотопного состава льда сопровождается увеличением средней концентрации сине-зеленых и зеленых водорослей. Более сильная активация наблюдается в воде с “облегченным” изотопным составом. Наибольший вклад в эту асимметрию вносят данные, полученные в июне, т.е. во время максимальной биологической активности, наблюдающейся не только в поверхностном слое, но и во всей толще льда. Можно было думать, что небольшие отклонения в изотопном составе воды в обе стороны от стандартного изотопного состава должны приводить к активации биологических систем, но это оказалось не так.
В 90-х годах появилась возможность проведения систематических лабораторных исследований с водой, обедненной дейтерием, полученной в промышленных масштабах. Первой работой в серии работ по биологическим изотопным эффектам воды, сильно обедненной дейтерием, следует, по-видимому, считать работу Г. Шомлаи [40]. В этой работе было показано, что использование воды 300 и 600 ppm вместо обычной воды, содержащей 150 ppm дейтерия, приводит к активации клеточной пролиферации в культуре фибробластов, а использование “облегченной” воды 30 ppm напротив к ее ингибированию. Рост культуры в воде 30 ppm характеризуется также существенно увеличенным лаг-периодом. Аналогичные результаты были получены для аденокарциномы, имплантированной в мышей, которых поили водой 30 ppm. Онкологическое направление исследований в Венгрии активно развивается и уже ряд лет ведутся клинические испытания. Выживаемость пациентов, принимавших длительное время только легкую воду 125 и 105 ppm совместно с обычными способами лечения, значительно выше, чем у пациентов, получавших лишь химиотерапию или лучевую терапию. Устраняются частично или полностью иммунодепрессивные эффекты, индуцированные лекарственными цитостатиками. Во всех случаях при употреблении облегченной воды наблюдается улучшение качества жизни и даже увеличивается продолжительность жизни пациентов [41-44]. Среди отечественных работ можно назвать обзор А.Тимакова, легко находимый в интернете [45].
Еще меньшие концентрации дейтерия в воде, вплоть до 4 ppm по дейтерию и 750 ppm по 18О (в контроле 140 и 1966 ppm соответственно), были исследованы и недавно представлены в работе [46]. Следует подчеркнуть, что эта вода была тщательно изучена и количество примесей в виде ионов Al, V, Cr, Mn, Ni, Fe, Cu, Zn, As, Cd, Pb во всех случаях не превышало 0,1 нМ. Температура плавления льда составила –1,50С, а температура кипения 93,70С. Это понижение температуры плавления даже превышает аналогичный эффект для 100 мМ раствора хлористого натрия или сахарозы. Такое сильное отклонение от закона Рауля и уменьшение на 7 градусов температуры кипения, по мнению авторов, свидетельствует о том, что легкая вода представляет собой новую ассоциированную жидкость не описанную ранее. Кинематическая вязкость уменьшена до 0,987 мм2/с (1,012 в контроле). Поверхностное натяжение, напротив, увеличено до 75,172 мН/м (72,860 в контроле). Очень большие изменения зарегистрированы для времени спин – спиновой релаксации протонов 0,35с (2,00 в контроле). В этой воде время жизни биоиндикатора Spirostomum ambigua оказалось в 10 раз меньше, чем в обычной воде. Такое же уменьшение зарегистрировано и в обогащенной дейтерием воде с 265 ppm.
Опишем результаты наших исследований с использованием легкой воды со следующим содержанием изотопов: D/H = 4 ppm, 18O/16O = 1335 ppm. Для достижения повышенной концентрации дейтерия использовали добавки D2O c концентрацией 99%. Исследована зависимость гидролитической активности Na,K-ATФазы из солевых желез утки от содержания дейтерия в воде. Активность фермента определяли методом Ратбуна и Бетлах по оптической плотности раствора. Предварительно была выполнена калибровка для воды с различным изотопным составом. Результаты приведены в табл. 2. Видно, что как уменьшение, так и увеличение концентрации дейтерия приводит к ингибированию активности фермента, выделенного из солевых желез утки.
|
Таблица 2. Активность белка Na, K-ATФазы. Указан доверительный интервал, соответствующий 5% уровню значимости |
|
|
Содержание дейтерия, % |
Активность, нМ/(час*мкг) |
|
0,0004 |
181 ± 4 |
|
0,0024 |
158 ± 6 |
|
0,015 |
179 ± 5 |
|
4,95 |
174 ± 4 |
|
9,9 |
169 ± 8 |
Результат, полученный в области повышенной концентрации дейтерия, противоположен относительно полученного нами ранее на препарате из мозга быка. По техническим причинам мы не смогли повторить эксперименты с препаратами из мозга быка. Активность фермента с минимальной концентрацией дейтерия не отличается от контрольного значения в обычной воде.
Проведены эксперименты с икринками вьюна Misgurnus fossilis. Икру оплодотворяли суспензией со сперматозоидами и помещали в раствор Гольтфретера, содержащий NaCl - 60 мМ, KCl - 0,7 мМ, CaCl2 – 0,9 мМ, NaHCO3 – 2,4 мМ и воду с исследуемым изотопным составом. Предварительно проверили, что оплодотворяющая способность сперматозоидов не зависит, среды, в которой размельчается семенник. После процедуры оплодотворения отбирали целые икринки и помещали в чашки Петри (не более 21 штуки на 1 чашку) с раствором изучаемого изотопного состава. Ход развития наблюдали в течение 6 суток с помощью микроскопа. Темпы развития икринок сравнивались с таблицами нормы развития икринок вьюна [47]. Аналогичные эксперименты проводили с икринками других особей. Суммарные данные приведены в табл.3.
|
Таблица 3. Развитие икринок вьюна. |
||||
|
|
исходное число икринок |
выжившие икринки к концу опыта (аномальные) |
выжившие, % |
аномальные/выжившие, % |
|
Контроль |
166 |
25 (6) |
15 |
24 |
|
Легкая вода |
165 |
53 (16) |
32 |
30 |
|
20% дейтерия |
40 |
0 |
0 |
0 |
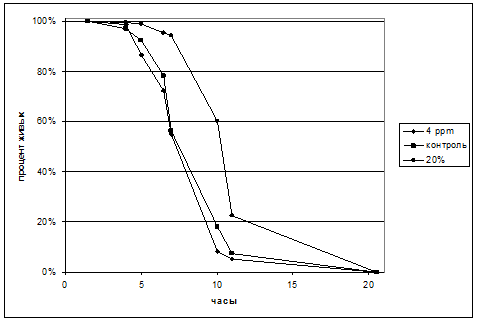
Из таблицы видно, что выживаемость в легкой воде 4 ppm вдвое превышает выживаемость в контроле. В воде с добавлением 20% дейтерия развития не происходило. Неоплодотворенные икринки, активированные водой, погибают через некоторое время. Можно проследить за динамикой этого процесса в воде с различным изотопным составом. Результат представлен на рис.8. Легкая вода не приводит к изменению кинетики выживаемости, но в воде, содержащей 20% дейтерия, процесс гибели клеток заметно заторможен.
|
|
|
Рис.8. Динамика гибели неоплодотворенных икринок. Погрешность составляет не более 3%.
|
Были проведены эксперименты с культивированием сперматозоидов человека в воде с разным изотопным составом. Сначала нативный эякулят центрифугировали, чтобы оставить только подвижные клетки. Суспензию со сперматозоидами помещали в пробирки с питательной средой изучаемого изотопного состава. Пробирки ставили в СО2 – инкубатор и культивировали при температуре 36,70С в течение 4-5 дней. В качестве критерия гибели использовался тест «мертвый-живой» с использованием набора Vital Screen. Результаты культивирования клеток в среде с содержанием дейтерия в интервале 5 – 40 % значимо не отличались между собой и показали 80 % гибель спустя сутки с начала эксперимента и не показаны на рисунке. Результаты, полученные при других концентрациях дейтерия, представлены на рис. 9.
|
|
|
Рис.9. Динамика гибели культивируемых сперматозоидов человека. Погрешность составляет 5%.
|
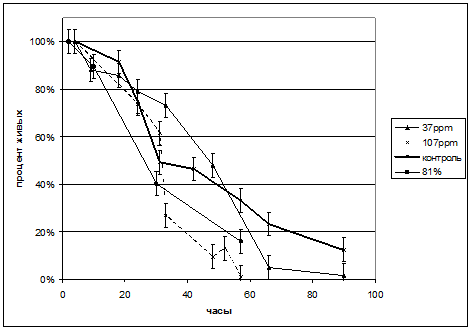
Видно, что преимущества в культивировании сперматозоидов человека в среде с пониженной концентрацией дейтерия не удается.
Исследовали также подвижность сперматозоидов, полученных из нативной спермы человека, замороженной спермы быка, нативных семенники вьюна и лягушки Hylarana temporalis. Сперматозоиды млекопитающих помещали в питательную среду TALP2, содержащей NaCl - 107,7 мМ, KCl - 3,1 мМ, Na2HPO4 - 0,36 мМ, CaCl2 -2 мМ, лактатдегидрогеназу - 25,4 мМ, пируватдегидрогеназу -1 мМ, гидроксиэтил-азинэтансульфокислоту (HEPES) - 10 мМ, NaHCO3 - 15 мМ, MgCl2 - 1 мМ. К сперматозоидам хладнокровных добавляли 0,7% раствор NaCl (в этом случае клетки неподвижны) и активировали их непосредственно перед измерением с помощью разбавления водой в 4 раза. Спермоанализатор регистрирует движение клеток и расчитывает среднее значение произведения концентрации
|
|
|
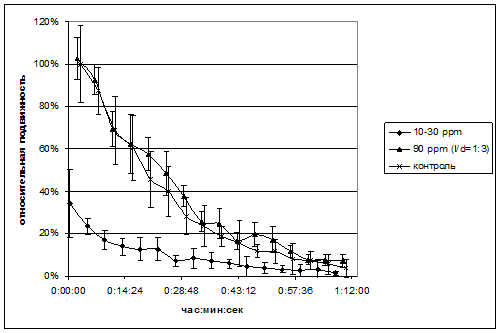
Рис.10. Зависимость подвижности сперматозоидов быка от времени. За начало отсчета по времени принято добавление замороженной спермы в питательную среду. Значения нормированы на первоначальное значение контроля. На графике отложено среднеквадратичное отклонение.
|
сперматозоидов на пройденное ими расстояние за определенный промежуток времени, которое будем называть подвижностью. Этот промежуток времени задается экспериментатором. Он составил 20 секунд для сперматозоидов млекопитающих и 10 секунд для хладнокровных. Концентрация клеток составляла 23±2 млн/мл для млекопитающих и 37±4 млн/мл для хладнокровных.
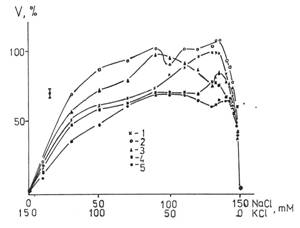
Сперматозоиды быка менее активны в средах с содержанием дейтерия от 13 до 50 ppm. В средах с облегченной водой начальные абсолютные значения были более чем в 2 раза меньше контрольных. При содержании дейтерия от 50 ppm до 90 ppm результат зависел от образца и мог не отличаться от контроля. Характер зависимости подвижности клеток от времени меняется от логарифмической (13-50 ppm) до экспоненциальной (≥90ppm). Результаты представлены на рис.10. Подвижность сперматозоидов человека представлена на рис.11.
|
|
|
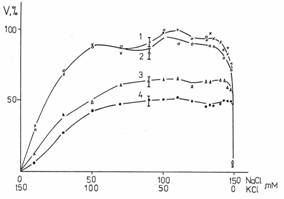
Рис.11. Зависимость подвижности сперматозоидов человека от времени. Время отсчитывается с первого измерения. На графике отложено среднеквадратичное отклонение.
|
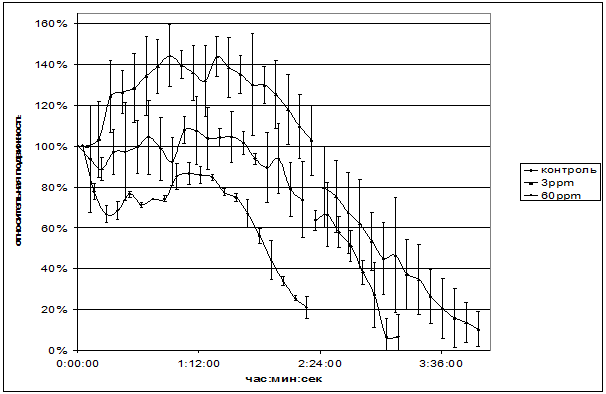
Подвижность сперматозоидов человека статистически значимо не отличалось от контроля в средах, содержащих 90±10 ppm дейтерия. При 30 ppm в течение 25 минут наблюдается 20% увеличение активности спустя 1 час после начала регистрации. Из рис. 11 видно, что в среде, содержащей 3 ppm дейтерия сначала даже наблюдается активация подвижности в течение часа, в то время как в обычной среде сначала происходит уменьшение активности.
Подвижность сперматозоидов лягушки и вьюна в среде, приготовленной на легкой воде с содержанием дейтерия 2,9 ppm, не отличалась от контроля. При концентрации дейтерия 370 ppm наблюдалось 3-х кратное увеличение подвижности сперматозоидов вьюна.
Наши последние исследования подтвердили, что зависимость изучаемых процессов от концентрации дейтерия в воде может иметь немонотонный характер, а уменьшение концентрации дейтерия относительно нормальной может приводить к аномально большим изотопным эффектам. Сами же эффекты, их величина и знак зависят от объекта исследования.
Причины влияния малых вариаций изотопного состава воды на биологические объекты в настоящее время не ясны. Родимов видел эту причину в изотопном замещении атома водорода в ДНК [30]. Но было показано, что эффекты малых концентраций дейтерия обнаруживаются и при исследовании выделенных ферментных систем [32, 33]. Мухачев В.М. обратил внимание на то, что концентрация дейтерия вполне сравнима с концентрацией основных микроэлементов, например концентрация атомов железа даже меньше, чем дейтерия [48]. Всем хорошо известно, какое большое значение имеют небольшие изменения содержания в живом организме железа, меди, кадмия, марганца, цинка и других микроэлементов, которых в десятки и сотни раз меньше, чем дейтерия. Однако, между ними существует принципиальное различие. Ионы металлов обладают очень высокими значениями констант связывания с соответствующими центрами макромолекул, а константа распределения дейтерия между водой и органическими молекулами менее десяти. Для объяснения наблюдаемых эффектов нужно найти центры связывания, обладающими константами селективности протонов порядка 103 − 10-4. Центры связывания протонов с такими характеристиками не известны и не возможны теоретически. Поэтому путь к объяснению рассматриваемых изотопных эффектов лежит в поиске высоко кооперативных больших систем, в которых замена одного атома протия на дейтрон может привести к изменению свойств всей системы в целом. Такими системами могут быть распределенные неравновесные системы, включающие мембраны, белки и воду и обеспечивающие пространственную и временную корреляцию событий, которые в иных условиях были бы редкими или вообще невозможными [49]. Изложенный материал убедительно подтверждает существование аномально больших изотопных эффектов в живых организмах и модельных системах при небольшой вариации изотопного состава водорода в воде. Наличие естественных природных изменений в соотношении изотопов воды может служить причиной некоторых экологических закономерностей, например, предпочтительного развития одного из вида микроорганизмов. Значение изотопных вариаций в природе, наряду с механизмами их действия на живые системы, еще предстоит выяснить.
Авторы благодарят Т.Н. Бурдейную и А.С. Чернопятко за предоставленную возможность работы с глубоко очищенной от дейтерия водой.
INFLUENCE OF ISOTOPIC CONTENT VARIATION OF WATER ON ITS BIOLOGICAL ACTIVITY
V.I.Lobyshev, A.A.Kirkina
Physics Department, M.V.Lomonosov Moscow State University. E-mail: lobyshev@yandex.ru
Литература
1. Lewis G.N. The biochemistry of water containing hydrogen isotope. // J. Amer. Chem. Soc., 1933, v.55, N8, p.3503-3504.
2. Taylor H.S., Swingle W.W., Eyring H., Frost A.A. The effect of water containing the isotope of hydrogen upon fresh water organisms. // J.Cell Comp. Physiol., 1933, v.4, N1, p.1-8.
3. Лобышев В.И., Калиниченко Л.П. Изотопные эффекты D2O в биологических системах. М., Наука, 1978, 213с.
4. Ферронский В.И., Поляков В.А. Изотопия гидросферы Земли. М., Научный мир, 2009, 632с.
5. Barnes T.C. A possible physiological effect of the heavy isotope of H in water/ // J. Amer. Chem. Soc., 1933, v.55, N10, p.4332-4333.
6. Barnes T.C. Futher observation on the physiological effect on the heavy hydrogen isotope on Spirogyra. // Amer. J. Botany, 1933, v.20, N10, p.181-182.
7. Barnes T.C., Larson E.J. Futher experiments on the physiological effect of heavy water and of ice water. // J. Amer. Chem. Soc., 1933, v.55, N 12, p.5059-5060.
8. Barnes T.C. The effect of heavy water of low concentration on Euglena. // Science, 1934, v.79, N 2050, p.370.
9. Barnes T.C. Alleged simulation of moulds by paraffin in heavy water. // Nature, 1934, v.134, N 3389, p.573-574.
10. Barnes T.C., Larson E.J. The effect of heavy water of low concentration on Spirogyra, Planaria and enzyme action. // Protoplasma, 1934, Bd.22, H.3, s.431-443.
11. Barnes T.C., Jahn T.L. Properties of water of biological interest. // Quart. Rev. Biol., 1934, v.9, N3, p.292-341.
12. Barnes T.C., Gaw H.Z. The chemical basis for some biological effects of heavy water. // J. Amer. Chem. Soc., 1935, v.57 N 3, p.590-591.
13. Richards O.W. The growth of yeast in water containing deuterium. // Amer. J. Botany, 1933, v. 20, N 10, p.679-680.
14. Richards O.W. The effect of deuterium on the growth of yeast. // J. Bacter., 1934, v.28, N3, p.289-294.
15. Mayer S.L.Alleged stimulation of moulds by paraffin in heavy water. // Nature, 1934, v.134, N3391, p.665.
16. Mayer S.L. Deuterium oxide and Aspergillus. // Science, 1934, v.79, N2044, p.210-211.
17. Macht D.I, Davis M.E. Some pharmacological experiments with deuterium. // J. Amer. Chem. Soc., 1934, v.56, N1, p.246.
18. Fox D.L.,Cupp E.E., McEwen G.F. Low concentration of deuterium oxide and the growth of marine diatoms.
1. // Proc. Soc. Expl. Biol. Med., 1936, v.34, N5, p.575-581.
19. Rea C.E., Yuster S. Effect of deuterium oxide on rat sarcoma R-39. // Proc. Soc. Expl. Biol. Med., 1934, v.39, N9, p.1058-1060.
20. Shoup C.S., Mayer S.L. The action of deuterium oxide in low concentration on the course of gas production by brewer’s yeast. // J. Tennessee Acad. Sci., 1935, v.10, N2, p.127-130.
21. Larson E.J., BarnesT.C. Parasition in heavy water of low concentration. // Nature, 1934,v.133, N3371, p.873-874.
22. Terao A., Inoue Y. Effect of the heavy water on the population growth of the water-flee, Moina macrocopa strauss. // Proc. Imp. Acad. (Tokyo), 1934, v.10, N8, p.513-514.
23. Brun J., Tronstad L. Some germination experiments with peas in heavy water. // K. Norske Vidensk. Selik. Fozhandl (Trondheim), 1935, v.7, (paper 48), p.171-173.
24. Curry J., Pratt R., Trelease S.F. Does dilute heavy water influence biological processes. //Science, 1935, v.81, p.275-277.
25. Weiser H.H. Influence of deuterium oxide on growth and morphology on lactobacilli. // Proc. Soc. Expl. Biol. Med., 1937, v.36, N2, p.151-152.
26. Fox D.L. Concerning enzymic reactions in heavy water. // J. Cell. Comp. Physiol., 1935, v.6, N3, p.405-424.
27. Ewart A.J. The influence of heavy water on plants. // J. Exp. Biol. Med., 1935, v.13, N1, p.9-15.
28. Родимов Б.Н. Снеговая вода – стимулятор роста и продуктивности животных и растений. // Сельское хоз. Сибири. Омск, 1961, №7б, с.66-69.
29. Родимов Б.Н., Маршунина А., Яфарова И. Действие снеговой воды на живые организмы.
2. // Сельскохозяйственное производство Сибири и Дальнего Востока. Омск, 1965, №4, с.56-57.
30. Торопцев И.В., Родимов Б.Н., Маршунина А.М., Яфарова И.О., Садовникова В.И., Лобина И.Г. Биологическая роль тяжлой воды в живых организмах. // Вопросы радиобиологии и гематологии. Изд. Томского ун-та, 1966, с.118-126.
31. Gleason J.D., Friedman I. Oats may grow better in water depleted in oxygen−18 and deuterium. //Nature, 1975, v.256, N5505, p.305.
32. Лобышев В.И., Твердислов В.А., Яковенко Л.В., Фогель Ю. Активирование Na,K-ATФазы малыми концентрациями D2О, ингибирование-большими. Биофизика 1978, N2, с. 390-391
33. Лобышев В.И., Фогель Ю., Яковенко Л.В., Резаева М.Н., Твердислов В.А. D2О как модификатор ионной специфичности Na,K-АТФазы. // Биофизика 1982, N4, с.595-603.
34. Ahmed K., Foster D. Studies on the effects of 2H2O on Na,K–ATPase. // Ann. N.Y. Acad. Sci., 1974, v.242,
3. p.280-291.
35. Лобышев В.И. Аномальное активирующее влияние тяжелой воды малой концентрации на регенерацию гидроидных полипов.// Биофизика 1983, N4, с.666-668.
36. Мельников И.А., Лобышев В.И. Фракционирование 18O в снежно-ледяном покрове центрального арктического бассейна. // Океанология 1985, N2, с.237-241.
37. Лобышев В.И., Мельников И.А., Есиков А.Д., Нечаев В.В. Исследование изотопного состава кислорода арктического дрейфующего льда в связи с проблемой активации роста микроорганизмов на границе тающего льда. // Биофизика, 1984, N5, с.835-839.
38. Есиков А.Д. Масс-спектроскопический анализ природных вод. М., Наука, 1980, 203с.
39. Николаев С.Д. Изотопно-кислородная палеотермия мирового океана в мезо-кайнозое. Состояние проблемы. В кн.: VII Всес. Симп. по стабильным изотопам в геохимии. М., изд. АН СССР, 1978, с.317-319.
40. Somlyai G., Janeso G., Jakli G., Vass K., Barna B., Lakies V., Gaal T. Naturally occurring deuterium is essential for the normal growth rate of cells. // FEBS, 1993, v.317, N1-2, p.1-4.
41. Somlyai G., Laskay G., Berkenyi T., Galbacs Z., Galbacs G., Kiss S.A., Jakli Gy., Jancso G. The biological effects of deuterium – depleted water, a possible new tool in cancer therapy. // Z. Oncol./J. of Oncol. 1998, v.30, N4, p.91-94.
42. Krempels K., Somlyai I., Somlyai G. A retrospective evaluation of the effects of deuterium depleted water consumption on 4 patients with brain metastases from lung cancer. // Integrative Cancer Therapies, 2008, v.2, N3, p.173-181.
43. Kovacs A., Guller I., Krempels K., Somlyai I., Janosi I., Gyongyi Z., Szabo I., Ember I., Somlyai G. Deuterium depletion may delay the progression of prostate cancer. // J. of Cancer Therapy, 2011, v.2, p. 548-556.
44. Avila.D.S., Somlyai G., Somlyai I., Aschner M. Anti-aging effects of deuterium depletion on Mn-induced toxicity in a C.elegans model. // Toxicology Letters, 2012, in press
45. Тимаков А.А. Основные эффекты легкой воды. // 8-ая Всерос. Конф. “Физико-химические процессы при селекции атомов и молекул. 6 – 10 ноября 2003.
46. Гончарук В.В., Лапшин В.Б., Бурдейная Т.Н., Плетенева Т.В., Чернопятко А.С. и др. Физико-химические свойства и биологическая активность воды, облегченной по тяжелым изотопам. // Химия и технология воды, 2011б т.33, №1, с.15-25.
47. Крыжановский С.Г. Эколого-морфологические закономерности развития карповых, вьюновых и сомовых рыб (Cyprinoidei и Suluroidei). // Труды Ин-та морф. животных, Изд. АН СССР, Вып. 1, с. 186-195.
48. Мухачев В.М. Живая вода. М., Наука, 1975, с.141.
49. Яковенко Л.В., Бутылин А.А., Твердислов В.А. Механические колебания и динамическая организация биомембран. // Биофизика, 1986, т.32, №6