На эту статью можно ссылаться, ее адрес в интернете: www.biophys.ru/archive/congress2009/pro-p87.htm
Орто/пара конверсия Н2О в воде и скачок «текучести» эритроцитов
через микрокапилляр при температуре 36.6±0.3 0С
С.М. Першин
Научный центр волновых исследований Института общей физики им. А.М.Прохорова РАН, 119991, Вавилова 38, Москва, pershin@orc.ru
PACS numbers: 42.62.Be;87.50.-a
Аннотация
Предложен механизм 0-100 % скачка «текучести» эритроцитов через микрокапилляр пипетки диаметром 1.3 мкм при температуре 36.4±0.3 0С, обнаруженного ранее [G.Artmann et al., Temperature Transitions of Protein Properties in Human Red Blood Cells, Biophysical Journal, 75 (1998) 3179]. Новый подход основан на экспериментальных фактах существования орто и пара спин-изомеров Н2О в воде, а также спин-селективном взаимодействии белков с пара-изомерами Н2О при формировании гидратных оболочек биомолекул [A.F. Bunkin, S.M. Pershin, A.A. Nurmatov, Laser Phys. Lett. 3(6), 275 (2006)]. Существенно, что образование гидратных оболочек белков и ДНК в водных растворах сопровождается увеличением Бриллюэновского сдвига до 0.4 cм-1 (в воде~0.25 cм-1), что указывает на образование в них льдоподобных структур (А.Ф.Бункин, С.М.Першин, Квант. Электроника, 39, 2009). Кроме этого, выявлен резонанс трансляционной энергии kT Броуновского движения и энергии вращательных квантов hΩ перехода 313-202 пара-изомеров Н2О при температуре 36.6 0С, который увеличивает вероятность возбуждения пара-Н2О при столкновениях. Столкновения смешивают квантовые состояния близко-расположенных уровней пара-Н2О (313, 285.2 см-1) и орто-Н2О (330, 285.4 см-1), соответственно, и индуцируют конверсию пара-изомеров в орто-Н2О. Предположено, что эта конверсия в льдоподобной гидратной оболочке гемоглобина (Hb) ускоряется при наличии в Hb кислорода и железа как катализаторов. Обосновано, что наблюдаемая потеря эритроцитом воды в капилляре обусловлена выходом через его мембрану орто-изомеров Н2О, которые освобождаются при «расплаве» гидратной оболочки Hb. «Расплав» способствует уплотнению молекул Hb и сопровождается увеличением плотности катализаторов, которые ускоряют конверсию и структурный переход гель-золь. Последовательность этих процессов подобна цепной реакции, которая запускает лавинно-образный скачок «текучести» эритроцитов через микрокапилляр. Этот же механизм позволяет обосновать наблюдаемое в работе [G.Artmann et al.] аномальное увеличение текучести водного раствора гемоглобина почти на порядок в области температуры 36.6 0С при повышении его концентрации в 1.7 раза. Рассмотренные здесь спин-спиновые взаимодействия относятся к сверхтонким взаимодействиям с типичной энергией 10-7 (kT) [Салихов К.М. 10 лекций по спиновой химии. - Казань: Казань: УНИПРЕСС, 2000, 143с.; http//chem.kstu.ru]. Несмотря на то, что это слабые взаимодействия, они управляют транспортом воды цитоплазмы через водные каналы клеточных мембран и проницаемостью эритроцитов через капилляры, снимая гипоксию либо поддерживая ее.
Ключевые слова: скачок «текучести» эритроцитов, особые температурные точки воды, орто-пара конверсия молекул Н2О.
Введение
Прохождение эритроцитов, доставляющих кислород ко всем органам, через капилляры имеет фундаментальное значение для обеспечения жизнедеятельности организма, особенно в экстремальных ситуациях и повышенных нагрузках, например, в спорте. В данной работе предпринята попытка обосновать роль сверхслабых взаимодействий, например, спин-спинового взаимодействия орто и пара-изомеров Н2О в управлении макроскопическими процессами такими как, доставка кислорода эритроцитом к периферийным отделам организма.
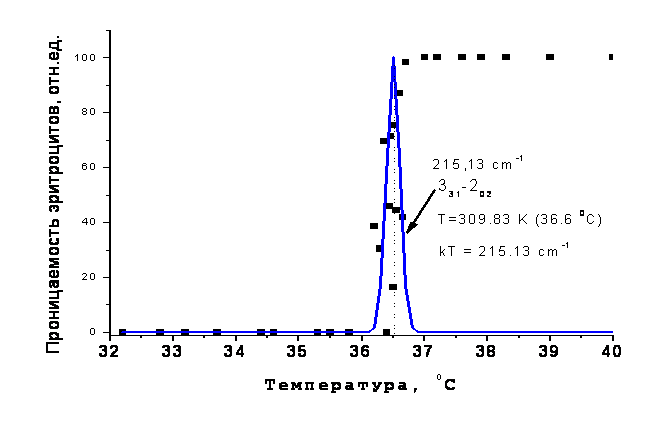
Декаду назад авторами работ [1, 2] был обнаружен скачок 0-100% текучести (по терминологии авторов) эритроцитов человека в капилляре при отборе пробы пипеткой с диаметром канала 1.3±0.2 мкм, обеспечивающей перепад давления 2.3 кПа. Наиболее удивительным было то, что скачок наблюдался в очень узком температурном диапазоне, 36.4±0.3 0С, совпадающем с нормальной температурой человека. Т.е. при температуре меньше 36 0С эритроциты не проходили в капилляр, а при температуре выше 37 0С – деформировались и протекали по капилляру с большой скоростью. На рисунке 1 показан характерный вид этой зависимости (по данным работы [1]).
|
|
|
Рис.1. Температурная зависимость проницаемости эритроцитов через микрокапилляр из работы [1]. |

Было установлено, что при температуре ниже скачка эритроцит частично втягивался в микрокапилляр, теряя при этом до 20 % воды, и оставался на конце пипетки, как упругий мячик. Иногда, в 45% случаев, при приближении к температуре скачка оболочка эритроцита разрывалась, что авторы относили к проявлению конформационных переходов в скелетообразующем белке мембраны- спектрине. Заметим, что диаметр эритроцита (около 7 мкм) существенно больше диаметра пипетки 1.3±0.2 мкм. При температурах выше скачка, эритроцит сжимался и втягивался в капилляр за несколько секунд, при этом объем эритроцита в капилляре уменьшался более чем вдвое. Это уменьшение сопровождалось потерей воды до 55% через водные каналы белка аквапорина в мембране эритроцита [1].
Кроме этого было установлено, что вязкость водного раствора гемоглобина, извлеченного из эритроцитов, уменьшается почти на порядок в окрестности этой же температуры при увеличении его концентрации в растворе в 1.7 раза, до 500 млг/мл. Такая концентрация гемоглобина, по оценке авторов [1], достигалась в сферической части эритроцита на конце пипетки при втягивании остальной части внутрь. Некоторые особенности с водными растворами биополимеров в окрестности этой температуры наблюдались и ранее (см. ссылки в работах [1, 2]), но не получили однозначной интерпретации.
За прошедшую декаду группа G.Artmann провела большое число различных экспериментов с целью выяснения природы этого скачка с использованием разных физических методов: ЯМР-спектроскопии, спектроскопии рассеяния, флюоресценции меток и белка спектрина, измерения дихроичности и ряд других. Однако механизм этого явления остается до конца невыясненным, несмотря на активное его изучение до настоящего времени [3-5]. В процессе исследования основное внимание было уделено изучению свойств гемоглобина (Hb) - основного белка в эритроцитах (92% сухого веса эритроцита составляет гемоглобин или до 330 млг/мл при нормальных условиях, что соответствует 0.25 части всего объема эритроцита). Предполагалось [3], что структурная перестройка Hb приводит к истончению гидратной оболочки белка Hb и выходу воды из эритроцита [4]. Была установлена обратимость потери воды: эритроциты принимали начальную форму и объем после возвращения в физиологический раствор.
Главный вопрос, почему скачок происходит при температуре 36.6 0С, оставался без каких-либо предположений. Кроме этого зарегистрированный «скачок» выхода молекул воды через мембрану эритроцитов от 20 до 55% в узком температурном диапазоне никак не соотносился авторами со свойствами воды [5].
Напротив, мы обратили на эти факты особое внимание, поскольку располагали некоторыми экспериментальными данными, опубликованными в наших работах [6-8], которые, на наш взгляд, прямо указывает на роль воды в этом явлении. В первую очередь, на определяющую роль воды указывает значение температуры скачка: при этой температуре у воды, как хорошо известно, наблюдается минимум теплоемкости при постоянном давлении. Кроме этого в ранних наших работах [6-8] мы обнаружили скачок центра ОН полосы в спектре комбинационного рассеяния в дистиллированной воде в окрестности температур 4, 19, 36-37 0С и других особых точках воды, в которых термодинамические параметры воды проходят через экстремум. Заметим, что температура 4 0С является определяющей для земноводных, а 19-20 0С – для хладокровных. Мы интерпретировали этот скачок как проявление фазового перехода второго рода, отражающего перестройку структуры сетки водородных связей в области температур особых точек, т.е. структуры воды [6-8]. Этот фазовый переход, как будет показано ниже, является определяющим в рассматриваемом явлении скачка эритроцитов.
Известно, что изучение механизма транспорта молекул воды через клеточные мембраны находится в области научных интересов многих лабораторий [9,10]. Цикл работ P.Agre с соавторами [9] был отмечен в 2003г. Нобелевской премией за открытие белка аквапорина и водных каналов в клеточных мембранах. Было установлено, что, несмотря на малый диаметр водного канала (3-3.8 Ангстрема), его пропускная способность одиночных молекул Н2О достигает несколько миллиардов за секунду. При этом механизм разрыва водородных связей (энергия связи ~3 ккал/моль) Н2О с соседними молекулами воды [9] перед входом в канал остался до конца невыясненным. В других ситуациях [10] проницаемость мембраны для молекул воды изучалась на моделях. Было показано, что проницаемость может быть обусловлена диффузией молекул воды. Результаты измерений температурной зависимости коэффициента самодиффузии через липидные бислойные мембраны также пока не имеет однозначного толкования [10]. Тем не менее, до сих пор не установлено, что оболочка эритроцита снабжена каким-либо температуро-зависимым клапанным механизмом, который бы открывал и закрывал водные каналы в окрестности температуры 36.6 0С и обеспечивал рассматриваемый здесь скачок проницаемости оболочки для молекул воды [1].
Заметим, что в цитируемых здесь и других работах наблюдаемые особенности свойств белков и воды обсуждаются без привлечения квантовых характеристик молекул Н2О, например, их отличия по ориентации спина протонов. Впервые [11], насколько нам известно, было установлено, что некоторые характеристики этих, сугубо квантовых, не имеющих классического аналога, спиновых изомеров, коррелируют с температурами особых точек льда и воды, например, 4, 20, 36-37 0С и др. Был выявлен резонанс значений тепловой энергии, kT, и значений энергии вращательных квантов, hΩ, близко-расположенных уровней орто и пара-изомеров молекулы Н2О. Кроме этого, как уже упоминалось выше, эти значения температуры известны нам не только как величины, при которых достигается экстремум термодинамических параметров: плотности воды (4 0С), поверхностного натяжения (20 0С) и теплоемкости (36.6 0С), но также наблюдается деформация огибающей и скачок центра ОН полосы [6-8] в спектроскопии комбинационного рассеяния.
Известно, что ядерные, орто и пара спин-изомеры молекулы Н2О [12,13], Н2 [14] и некоторых других молекул отличаются взаимной ориентацией спина протонов 1Н. Орто-изомер имеет параллельно ориентированные спины (суммарный магнитный момент равен единице, J=1). Напротив, суммарный магнитный момент пара-изомера равен нулю, J=0, из-за противоположно ориентированных спинов протонов. Поэтому равновесное орто/пара отношение согласно квантовой статистике равно 3:1, например, в воздухе при комнатной температуре [11-14]. Существенно, что часть пара-изомеров Н2О при этой температуре находится на основном уровне и не вращается, в то время как основной вращательный уровень орто-изомеров смещен на 23.8 см-1 от нулевого уровня (что соответствует энергии ~34 К на температурной шкале) и поэтому свободные орто-молекулы вращаются всегда пока заселен этот и выше лежащие уровени [15].
Орто/пара конверсия в отдельной молекуле строго запрещена в дипольном приближении [14]. Например, жидкий пара-водород может сохраняться несколько месяцев с медленным спонтанным обогащением орто-изомерами, механизм которого пока до конца неясен [16]. Однако присутствие катализаторов в виде парамагнитных примесей, таких как, кислород, железо, медь или других переходных металлов, существенно увеличивает скорость орто-пара конверсии [16,17].
Предметом настоящей работы является обоснование гипотезы о квантовой природе 0-100 % скачка протекания эритроцитов через микрокапилляр пипетки в окрестности температуры 36.4 0С на основе экспериментальных данных о наблюдении специфических свойств орто-пара спин-изомеров Н2О в воде и, в так называемой, биологической воде- гидратных оболочках биополимеров и водных растворах белков, ДНК и др.
Экспериментальные данные
Методом поляризационной четырехфотонной спектроскопии движения молекул Н2О в воде и водных растворах биополимеров в ТГц и субтерагерцовой области спектра [18-20] нам впервые удалось обнаружить резонансные линии, частоты которых совпадали с частотами вращательных переходов свободных молекул орто-пара спин-изомеров Н2О [15]. Более того, было установлено, что в растворах белков и ДНК наблюдалось селективное взаимодействие биополимеров с пара-молекулами Н2О [18, 19]. На рисунке 2 представлено два спектра четырехфотонного рассеяния в воде (круги) и водном растворе белка (квадраты) в терагерцовой области частот (2-3 ТГц) из нашей работы [20].
|
|
|
Рис.2. Четырехфотонный спектр воды (круги) и водного раствора белка (квадраты) [20]; орто-резонансы (тонкие стрелки), пара-резонансы (толстые стрелки). |

Из рисунка видно, что спин-селективное взаимодействие проявлялось в уменьшении амплитуды линий пара-молекул (79.8 см-1) в спектре (квадраты), которое интерпретировалось нами как формирование водородосвязанных гидратных оболочек белков из пара-изомеров молекул Н2О [20]. Часть пара-изомеров Н2О (в соответствии с Больцмановским распределением населенностей по энергетическим уровням) могут не вращаться в основном состоянии при комнатной температуре и температуре 36.6 0С [15]. По этой причине эти молекулы могут «захватываться» из контактного слоя водного раствора с большей вероятностью, поскольку они проявляют большую адгезию к гидрофильным остаткам биополимерных молекул, чем орто-молекулы. Увеличение сечения взаимодействия пара-изомеров Н2О с биополимерами было экспериментально обосновано в работе [21] при распространении паров Н2О в контейнере над поверхностью сухой пленки с биомолекулами и обогащении паров воды орто-изомерами на выходе контейнера. В растворах нам удалось обнаружить спин-селективное взаимодействие пара-Н2О с белками и ДНК, насколько нам известно, впервые [20]. Такое взаимодействие может приводить к формированию структурированного слоя воды около молекул белка. Примером может служить белок куриного яйца, который подобен гелю и стремится, скорее, к сохранению объема после разрушения скорлупы, а не растекается тонкой пленкой как водный раствор, например, поваренной соли. Другим примером является желе (судень) из водного раствора желатина (производная белка коллагена), льдоподобная форма которого сохраняется при положительных температурах.
Существенно, что наши результаты и выводы [18-20] о свободном вращении молекул в объемной воде и вблизи больших молекул биополимеров на основании прямого наблюдения резонансных линий вращательных переходов согласуются с данными работ группы Нобелевского лауреата A.H. Zewail о вращательной диффузии молекул Н2О в так называемой, биологической воде [22, 23].
Весьма важным свидетельством о специфических свойствах гидратного слоя на контактной границе с полимерным гелем являются экспериментальные данные G. Pollack [24, 25]. В изящном эксперименте со взвешанными в воде наносферами Поллак показал, что формирование контактного гидратного слоя на поверхности геля сопровождается полным вытеснением наносфер из этого слоя. При этом толщина слоя увеличивалась со временем до сотен микрон. Автор считает, что этот слой может быть образован структурированной водой с одинаково ориентированными диполями молекулы Н2О [24]. В монографии [25] автор убедительно показывает на основе многочисленных экспериментальных данных ряда лабораторий, что гидратный слой биомолекул состоит из структурированной воды. Этот слой придает всей конструкции биополимера определенную жесткость и упругость. При этом разрушение структуры этого слоя сопряжено с конформационным переходом биомолекулы. Однако, несмотря на возможность существования структуры в гидратном слое воды, неясно, каким образом подобный комплекс белок-вода будет чувствителен к фиксированной температуре, например, 36.6 0С.
Новые экспериментальные данные, полученные нами и другими авторами в последние годы, позволяют предположить, что гидратный слой имеет льдоподобную структуру сетки водородных связей. Тогда можно обосновать существование особых температурных точек структурных переходов, значение которых совпадает с известными специфическими температурами для воды [11]. Сравнительно недавно [26] было показано, что гидратный слой, так называемых, антифризных белков (хитин у насекомых) имеет гексагональную структуру льда Ih, что позволяет насекомым переносить зимнюю спячку при низких температурах без разрушения тканей (об этом упоминает и Поллак [25]). Приняв этот факт во внимание и, зная, что при кристаллизации воды лед вытесняет все примеси на грань кристалла, можно объяснить вытеснение наносфер из гидратного слоя, которое наблюдал Поллак [24], формированием льдоподобной структуры воды в этом слое.
Более того [27], нам удалось обнаружить, что ОН полоса комбинационного рассеяния в дистиллированной воде комнатной температуры содержит спектральную компоненту (3220 см-1) массивного льда. В последующем эксперименте [28] при повышении температуры воды было установлено, что амплитуда этой компоненты уменьшается, но остается обнаружимой в воде вплоть до температуры кипения. Рис. 3 показывает эту температурную эволюцию огибающей ОН полосы после вычитания вклада обобщенного среднего [27]. Из рисунка также видно, что с повышением температуры увеличивается амплитуда (вклад) спектральных компонент 3450 и 3600 см-1, которые являются характеристическими частотами тетрамеров и димеров, соответственно.
|
|
|
Рис.3. Температурная эволюция огибающей ОН полосы валентных колебаний молекул воды |

Таким образом, в жидкой воде существуют устойчивые молекулярные комплексы с характерными частотами массивного льда, тетрамерных и димерных кластеров наряду с короткоживущими комплексами, которые формируют гладкую огибающую обобщенного среднего. Ранее нами [29] было обнаружено, что гравитационный центр ОН полосы воды совершает флуктуационные гармонические осцилляции около среднего положения. При этом суммирование спектров со смещением центра в низкочастотную область дает огибающую ОН полосы, которая подобна спектру массивного льда. Фактически этими спектральными данными нам впервые удалось подтвердить гипотезу о существовании устойчивой динамической льдоподобной («айсберги») структуры сетки водородных связей воды. Этот вывод согласуется также с измерением в воде двух значений скорости звука, одно из которых (~1590 м с-1) характерно для жидкой воды, а другое – (~3200 м с-1) для массивного льда [30]. Более того, сравнительно недавно [31] была получена пленка массивного льда при комнатной температуре на графитовой подложке в градиентном поле иглы атомно-силового микроскопа.
Далее было установлено [20], что формирование гидратного слоя белков и других биополимеров в водных растворах сопровождается селективным взаимодействием с пара-изомерами Н2О (см. рис.2). Недавно [32] нами было обнаружено, что Бриллюэновский сдвиг, который пропорционален скорости звука, в таких растворах больше, чем в воде и соответствует сдвигу во льдах. Ранее [33] было измерено, что спектральная компонента (~3220 см-1) ОН полосы массивного льда доминирует в биокристаллах белка лизоцима и тауматина. Причем было установлено, чем совершеннее кристалл, тем больше вклад спектральной компоненты льдоподобной сетки водородных связей.
Совокупность изложенных экспериментальных фактов показывает, что вода в гидратных слоях биомолекул, скорее всего, структурирована и имеет структуру льда. Эксперименты G.Pollack дают основание заключить, что гидратная оболочка биомолекул может обладать определенной упругостью, например, упругое состояние желе из раствора коллагена при комнатной температуре [24, 25]. Жесткость льдоподобной структуры оболочки поддерживает форму белка (эритроцитов) и препятствует его деформации (сворачиванию в клубок с минимальной поверхностной энергией в соответствии с термодинамикой). Более того, спин-селективное взаимодействие (см. рис.2) пара-изомеров Н2О с белками в водных растворах [20] позволяет ожидать, что гидратная оболочка образована преимущественно пара-изомерами. Недавно [34] нами было установлено, что равновесное отношение орто/пара 3:1, справедливое для газов при комнатной температуре, смещается в воде в сторону увеличения числа пара-изомеров Н2О более чем в 2 раза. Таким образом, было показано, что вода неравновесная, в смысле спиновой температуры, жидкость. Поэтому вода в таком состоянии способна к изменению этого соотношения в сторону равновесного, т.е. увеличения числа пара-изомеров, особенно при повышении температуры [34].
Квантовая природа скачка проницаемости эритроцитов через микрокапилляр
Принимая во внимание изложенные выше экспериментальные факты, рассмотрим физический механизм наблюдаемого скачка проницаемости эритроцитов в окрестности температуры 36.6 0С. Учтем, что спин-спиновые [35] и магнитные взаимодействия [36] индуцируют конверсию орто и пара-изомеров при столкновениях при наличии катализаторов (железо, медь, кислород и др) [16, 17, 37]. Ключевым фактором для обоснования предлагаемого механизма скачка здесь является обнаруженный нами ранее [11] факт совпадения энергии квантов (hΩ) вращательных переходов изомеров Н2О и тепловой энергии kT в окрестности 36-37 0С. Анализ вращательных спектров молекулы Н2О [15] показал, что в области этой особой точки воды существует резонанс энергий:
энергия кванта hΩ [(331-202) = 215.13 см-1 ] = энергии kT(при T=309.83 K=36.6 0С) (1)
Рис.4 демонстрирует взаимное положение модельного контура резонансной линии перехода 331-202 пара-изомера (сумма нижних индексов четная) Н2О с полушириной 0.4 см-1, которая сопоставима с экспериментальным откликом (см. рис.2), и экспериментальных точек скачка из работы G. Artmann [1]. Как видно из рисунка совпадение резонансной частоты перехода с энергией теплового движения оказалось лучше, чем ожидалось. Т.о. нам удалось найти один из базовых параметров эксперимента, а именно, значение температуры скачка «текучести» эритроцитов, которое определяется энергией кванта вращательного перехода молекулы воды. Остается вопрос, как формируется лавино-образной характер скачка в узком диапазоне температур при большой ширине функции Максвелла распределения молекул по тепловым скоростям [11]?
Для ответа на этот вопрос необходимо принять во внимание другое аномальное явление, которое наблюдал G. Artmann [1] при температуре 36.6 0С: снижение вязкости водного раствора гемоглобина почти на порядок при повышении его концентрации в 1.7 раза. Кроме этого напомним о существовании смешанных квантовых состояний [37, 38] близко-расположенных возбужденных уровней пара-изомера 331 (285.219 см-1) и орто-изомера 330 (285.419 см-1) в процессе резонансного столкновения [38 и ссылки в ней]. Подобный обмен энергией возбуждения, индуцированный столкновениями, хорошо известен по схеме накачки гелий-неонового (He-Ne) лазера [39]. В He-Ne лазере атомы гелия из основного состояния 11S переводятся (при столкновениях с электронами газового разряда) в возбужденные состояния 23S и 21S. Эти атомы гелия передают энергию возбуждения атомам неона и переводят их в состояния 4S и 5S, несмотря на некоторую отстройку от точного резонанса энергий этих уровней.
В нашем случае может происходить конверсия спинового состояния одной и той же молекулы Н2О, электродипольный запрет на которую снимается в возбужденном состоянии молекулы в неоднородном магнитном поле катализатора [14,16,17,35-37]. Заметим, что расстройка орто-пара резонансных уровней (0.2 см-1) может компенсироваться (как и в случае He-Ne лазера) за счет энергии столкновений. Из Рис.4 видно, что 100% проницаемость эритроцитов достигается в окрестности температуры ~36.9 0С. Это значение отличается от резонансного (hΩ (331-202) = 215.13 см-1) на ~0.3 0С, что согласуется с величиной расстройки резонансов, поскольку известно, что 1 0С ≈ 0.69434 см-1 [11] и тогда получим:
0.2 см-1 = 0.2/0.69434 ≈ 0.29 0С (2)
Действительно авторы [1] отмечают сокращение времени сжатия оболочки эритроцита на входе в пипетку при повышении температуры раствора в этом диапазоне. То есть максимальное значение на кривой рис.1 достигается тогда, когда максимум наиболее вероятной скорости молекул соответствует температуре 36.9 – 37 0С.
Рассмотренные факты показывают,
что в окрестности резонансной температуры наиболее вероятная скорость молекул
определяется величиной kT в соответствии с распределением
Максвелла, поэтому часть молекул пара-Н2О после столкновения и
резонансного обмена окажется в состоянии орто-изомера
на уровне 330 (285.419 см-1). Такой переход приведет к
уменьшению концентрации пара-изомеров, к разрыхлению гидратной оболочки и снижению ее упругости. Поскольку
вероятность конверсии увеличивается в поле катализатора [35-38], очевидно, что
максимальная скорость конверсии будет достигаться в слое, прилегающем к
молекуле гемоглобина, который содержит молекулы-катализаторы, кислород и
железо. Предложенный механизм «расплава», разрыхления оболочки, уменьшение ее
толщины и появление «текучей» фракции в воде в виде орто-изомеров
подтверждается экспериментальным фактом аномального снижения вязкости раствора
гемоглобина при температуре 36.6
В результате развивается самоподдерживающаяся последовательность процессов типа цепной реакции: (1)-в окрестности 36.6 0С увеличивается вероятность конверсии пара- Н2О в орто-Н2О в гидратном слое около белка гемоглобина; (2)-гидратный слой около Hb истончается («плавится») и фрагментарно разрушается его льдоподобная структура, которая препятствовала слипанию молекул Hb, например, в «монетные» столбики; (3)-молекулы гемоглобина сближаются друг с другом (под действием давления пипетки); (4)-объем эритроцита уменьшается почти на 55% из-за выхода освободившихся при конверсии орто-изомеров Н2О из оболочки; (5)-концентрация катализаторов (железо и кислород в Hb) в единице объема увеличивается; (6)-конверсия пара-орто Н2О ускоряется [16, 17, 31-33], формируя лавинно-образный скачок: 1-2-3-4-5-6-1.
Заметим, что при частичном втягивании оболочки эритроцита в пипетку и потери до 20% воды концентрация гемоглобина в сферической части эритроцита на конце пипетки увеличивается до величины 500 млг/мл. Орто-молекулы Н2О, образованные после столкновений и конверсии будут находиться в возбужденном состоянии на уровне 330 (285.419 см-1), т.е. вращаться интенсивнее, чем при комнатной температуре (~202 см-1). Это увеличит вероятность выхода орто-Н2О из эритроцита поодиночке через водные каналы аквапорина в мембране [9]. Напомним, что в эксперименте [1] наблюдается скачок выхода до 55% воды из эритроцита при его проникновении в капилляр пипетки. Повышение концентрации орто-изомеров с одновременным разрыхлением гидратной оболочки молекулы гемоглобина объясняет аномалию уменьшения вязкости водного раствора гемоглобина в окрестности температуры 37 0С [1].
Наличие скелетообразующего белка спектрина в оболочке эритроцита и способность белков селективно взаимодействовать (см. рис.2) с пара-молекулами Н2О [20] позволяет предположить, что рассмотренный выше механизм проявляется и в наблюдаемом [1] явлении разрыва оболочки эритроцита.
Механизм разрыва оболочки эритроцита
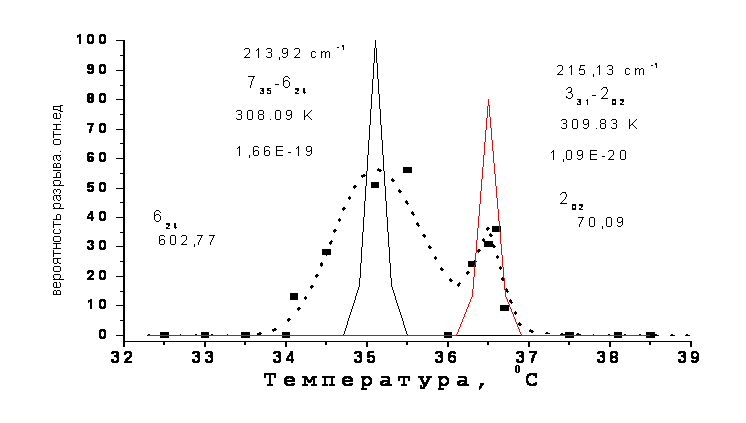
Во время изучения температурной зависимости скачка проницаемости эритроцитов через микрокапилляр авторами работы [1] было обнаружено, что иногда (с вероятностью до 50%) оболочка эритроцита разрывалась, когда он находился на кончике пипетки. Было установлено, что температурная зависимость вероятности разрыва имеет характерный резонансный вид в окрестности температуры 35 0С. На Рис. 5 показаны экспериментальные точки (квадраты) этой зависимости из работы [1], которые аппроксимировались одной огибающей с максимумом при 35 0С. Авторы работы предполагают, что это явление обусловлено конформационным переходом белка спектрина без какого-либо обоснования значения температуры резонансной зависимости.
В отличие от авторов [1] мы аппроксимировали эти данные двумя перекрывающимися огибающими (пунктир) гауссовской формы с температурой максимума 35 и 36.6 0С, соответственно. Полученное совпадение нам представляется более оптимальным, чем при аппроксимации одной огибающей [1]. Причем различие в амплитуде и ширине огибающих указывает на некоторое отличие механизма разрыва оболочки при температуре 35 и 36.6 0С. Так узкий пик, совпадающий по температуре и ширине с резонансной зависимостью скачка проницаемости эритроцитов (рис.4), скорее всего, обусловлен процессом с участием резонансного обмена энергией kT ≈ hΩ (1).
|
|
|
Рис.4. Скачок проницаемости эритроцитов (квадраты) и модельный контур линии квантового перехода hΩ (331-202) = 215.13 см-1 пара-изомера Н2О, построенный с полушириной 0.4 см-1 на температурной шкале. |
Анализ вращательного спектра пара-Н2О показывает, что помимо описанного выше резонанса при температуре 36.6 0С (1) имеется резонанс в окрестности 35 0С с энергией вращательного кванта на переходе 735-624 пара-изомера Н2О:
энергия кванта hΩ [(735-624) = 213.92 см-1] = энергии kT(при T= 308.09 K≈35 0С) (3)
Однако, в отличие от резонанса (331-202) при температуре 36.6 0С, здесь в окрестности верхнего уровня 735 перехода (735-624) нет близко–расположенного уровня орто-изомера. Этот фактор, скорее всего, приводит к уширению температурной зависимости на рис.5 в области 35 0С и сдвигу скачка проницаемости эритроцитов в область 36.6 0С, где образуются смешанные состояния орто-пара изомеров Н2О при столкновениях и вероятность орто-пара конверсии возрастает из-за наличия близко-расположенных уровней 330 и 331.
|
|
|
Рис.5. Температурная зависимость вероятности разрыва оболочки эритроцита (квадраты и пунктир) по работе [1] и линии вращательных переходов 735-624 (35 0С) и 331-202 (36.6 0С) пара-Н2О. |
Далее для понимания механизма разрыва оболочки эритроцита обратимся к экспериментальным результатам серии работ Захарова с соавторами [40-42], который изучал светокислородный эффект, температурные изменения показателя преломления водных растворов и суспензии эритроцитов при их концентрации 0.1 – 1.5 %, а также деформацию оболочки эритроцитов при вариации температуры от 15 до 40 0С. Особое внимание было уделено взаимосвязи структурных изменений водной среды и белков вне и внутри клеток, а также влиянию синглетного и триплетного кислорода на их свойства.
Было установлено, что оба параметра, деформация эритроцитов и показатель преломления внеклеточной водной суспензии изменяются синхронно, достигают экстремума в области 36.6 0С и при других значениях температуры. Природа этих значений температуры не обосновывалась. Авторы предположили, что наблюдаемые изменения отражают перестройку структурных форм гидратных слоев гемоглобина внутри эритроцита, а также снаружи оболочки, которая содержит скелето-образующий белок спектрин.
Наиболее интересным является регистрация уменьшения показателя преломления внеклеточного водного раствора при появлении в нем эритроцитов и увеличении их концентрации (см. рис. 10 и 8 в [42]). Учитывая, что показатель преломления льда (~1.309) меньше, чем у воды (~1.333), авторы интерпретируют наблюдаемое снижение показателя преломления как образование льдоподобных структур, типа гидрогеля, в гидратном слое оболочки эритроцита. Оценка показала, что толщина гидрогеля, прилегающего к мембране эритроцита, сравнима с диаметром самого эритроцита и составляет 5-10 мкм. Авторы считают, что такие гидратные оболочки метастабильны, существуют как у эритроцитов, так и альбуминов в определенных температурных интервалах. Следуя этой логике, отнесем сюда и гидратные оболочки гемоглобина. Предполагается, что в температурных интервалах с пониженным значением показателя преломления раствора и деформируемости эритроцитов происходят фазовые переходы в биомембранах и конформационные переходы в белках [42]. Предполагается также, что существенное влияние на динамическую микроструктуру водной среды оказывает молекулярный кислород, поскольку его удаление из воды приводит к исчезновению наблюдаемых особенностей.
Особое внимание авторы уделяют возможному проявлению магнитного взаимодействия между парамагнетиком О2 в триплетном состоянии и ядерными моментами протонов Н2О. При этом они допускают, что «магнитные силы неспаренных электронов спинов кислорода могли бы оказать ориентирующее влияние на спины протонов в кластере, однако этому препятствуют тепловые флуктуации. Такое влияние невозможно объяснить с позиции статического равновесия, т.к. в нормальных условиях при T=300 К
μВ B ~ 10-7 kT (4)
где μВ – магнетон Бора, B≈0.5 Гс – индукция земного поля».
Действительно, способность к
изменению свойств воды (уменьшается краевой угол капли на поверхности металлов)
после обработки в сильном (3-11 Тесла) поле постоянного магнита только при
наличии кислорода экспериментально обоснована в работе [43, 44]. Установлено,
что удаление растворенного кислорода или воздуха из воды делают ее
нечувствительной к магнитному воздействию. Существенно, что при добавлении
солей-электролитов, таких как, NaCl, KCl и CaCl2 с концентрацией раствора до
Обнаруженные нами орто и пара спин-изомеры Н2О в воде [18-20] дают нам основание для разработки нового подхода к пониманию наблюдаемых явлений в омагниченной воде также. Так в предлагаемом нами механизме тепловые флуктуации и столкновения обеспечивают и поддерживают спин-конверсию орто-пара Н2О в магнитном поле парамагнетика О2 как катализатора (см., например, [16, 17, 37]). Поэтому удаление кислорода из воды будет уменьшать вероятность орто-пара конверсии. Возможно, именно этот фактор проявлялся в работах [42-44] как нечувствительность воды без кислорода к магнитному воздействию. Напротив, известно, что повышение концентрации триплетного кислорода приводит к увеличению скорости орто-пара Н2О конверсии, которое было недавно экспериментально обосновано в работе Xavier Michaut et al. [37].
Тогда наблюдаемый иногда разрыв оболочки эритроцитов (рис.5) в области «язычка» (наиболее напряженной части мембраны) в микрокапилляре [1] может быть обусловлен пара-орто конверсией, которая может происходить одновременно в гидратном слое снаружи эритроцита, а также внутри его у молекул гемоглобина. Появление «свободных» орто молекул Н2О внутри эритроцита, не способных быстро выйти через мембранные каналы аквапорина и перемещающихся в «язычок» оболочки, формируют гидроудар из-за перепада давления в микрокапилляре. Вероятность разрыва оболочки под действием такого гидроудара при температуре 36.6 0С меньше чем при 35 0С (рис.4), поскольку скачок деформируемости оболочки эритроцита обеспечивает его втягивание в микрокапилляр без разрушения (см. выше конкурирующий процесс (3)).
Выводы
Таким образом, в воде выявлен резонанс энергии вращательного кванта hΩ на переходе 331-202 пара-изомера Н2О с энергией Броуновского движения kT при температуре 36.6 0С. Наличие такого резонанса увеличивает вероятность пара-орто конверсии при столкновениях и образовании смешанных квантовых состояний близко-расположенных уровней орто (330) и пара-изомера (331). Сформулирована и обоснована гипотеза о льдоподобной структуре водородных связей в гидратных оболочках белков и эритроцита, которые формируются преимущественно из пара-изомеров молекул воды [45]. Предложена квантовая природа и рассмотрен физический механизм наблюдаемого в работе [1] 0-100% скачка проницаемости эритроцитов через микрокапилляр при температуре 36.6 0С, который учитывает сугубо квантовые свойства молекул воды, отличающихся взаимной ориентацией спина протона. В соответствии с этим механизмом в цитоплазме эритроцита развивается самоподдерживающаяся последовательность процессов типа цепной реакции при температуре 36.6 0С: (1)-в окрестности 36.6 0С увеличивается вероятность конверсии пара- Н2О в орто-Н2О в гидратном слое около белка гемоглобина; (2)-гидратный слой около Hb истончается («плавится») и фрагментарно разрушается его льдоподобная структура, которая препятствовала слипанию молекул Hb, например, в «монетные» столбики; (3)-молекулы гемоглобина сближаются друг с другом (под действием давления пипетки); (4)-объем эритроцита уменьшается почти на 55% из-за выхода освободившихся при конверсии орто-изомеров Н2О из оболочки; (5)-концентрация катализаторов (железо и кислород в Hb) в единице объема увеличивается; (6)-конверсия пара-орто Н2О ускоряется [16, 17, 35-37], формируя лавинно-образный скачок: 1-2-3-4-5-6-1.
Этот же механизм проявляется в аномальном (почти на порядок) увеличении текучести водного раствора молекул гемоглобина при повышении (!!) его концентрации в 1.7 раза. При этом, в отличие от рассмотренной выше цепочке процессов, здесь молекулы орто-Н2О остаются в растворе и способствуют уменьшению вязкости, поскольку в свободном состоянии они всегда вращаются [15].
Изложенные факты дают основание предположить, что подобный механизм освобождения единичных молекул из сетки водородных связей возможен вблизи водных каналов белков аквапоринов [9, 46]. В работе [9] было установлено, что транспорт молекул воды через клеточные мембраны происходит по одной молекуле со скоростью до 3 109 молекул в секунду. Однако механизм разрыва водородных связей и освобождения молекул остается пока неустановленным.
Практическое применение понимания работы механизма скачка проницаемости эритроцитов через микрокапилляры при температуре 36.6 0С более актуально. Каждый из нас встречался и будет иметь дело с образованием внутритканевой гематомы после ушиба или травмы связок и пр. Первая помощь- приложить охлаждающий компресс. При этом действие этого компресса обосновывают сужением капилляров и сосудов, которое уменьшает вероятность вытекания крови из разорванных капилляров и поэтому стараются применить глубокое охлаждение, которое одновременно дает эффект анестезии. К этому механизму сужения сосудов надо добавить рассмотренный выше механизм непроницаемости микрокапилляров для эритроцитов при охлаждении участка ткани всего на 1-2 градуса от значения 36.6 0С (см.рис.1). Т.е. не надо тратить время на поиски источника глубокого охлаждения (пары жидкого азота, например), а быстро воспользоваться любым предметом комнатной температуры с большой теплоемкостью (металл: ложка, монеты или камень и пр.) для локального снижения температуры области травмы на 1-2 градуса. Главное здесь – потеря времени усугубляет последствия травмы.
Работа выполнена при финансовой
поддержке Российского фонда фундаментальных исследований, грантов №
05-02-16020, № 08-02-00008 и №09-02-01173, а также гранта Президента РФ
государственной поддержки ведущих научных школ РФ № НШ-8108. 2006.2
СПИСОК ЛИТЕРАТУРЫ
1.
Artmann G.M.,
Kelemen C., Porst D., Buldt
G., and Chien S. // Biophys.
J., 1998, 75, 3179.
2.
Artmann
G.M., Sung K.-L. P., Horn T., Whittemore D., Norwich
G., and Chien S.// Micropipette aspiration of human
erythrocytes induces echinocytes via membrane phospholipid translocation. Biophys.
J. 1997, 72, 1434–1441.
3.
Kelemen C., Chien
S., and Artmann G.M.//Biophys.
J. 2001, 80, 2622.
4.
Digel I., Maggakis-Klemen Ch., Zerlin
K.F., Kasischke N., Kayser P., Porst D., Artmann A.T., and Artmann G.M.// Biophys.
J. 2006, 91, 3014.
5.
Stadler A.M., Digel I., Artmann
G.M., Embs J.P., Zaccai G., and Buldt G. // Biophys. J.
BioFast, August 2008, http//www.biophysj.org
6.
Pershin
S. “The model of the temperature shifting and jumping of the Spontaneous Raman
Scattering O-H bands statistical center in water // Physics of Vibrations,
1997, 61, 4.
7.
Bunkin
A.F. and Pershin S.M. Temperature anomalies of liquid
water stretching vibrations Raman band envelope // Physics of Vibrations,
1997, V.61(3), 158-164; Першин С.М., Бункин А.Ф. “Скачок” центра и ширины огибающей спектральной полосы КР валентных колебаний ОН при фазовых переходах первого и второго рода в воде // Оптика и Спектроскопия,
1998, 85(2), 190-193.
8.
Bunkin
A.F., Lyakhov G.A., Suysov
N.A., and Pershin S.M. Temperature Anomalies in Water
Raman Spectrum: Identification of phase Transitions in Hydrogen Bond Network //
Physics of Vibrations. 1999. 7(2),75-80; Bunkin
A.F., Lyakhov G.A., Suysov
N.A., and Pershin S.M. Sequence of water
thermodynamic singularities in Raman spectra // J. Raman Spectrosc.
2000. 31. 857-861.
9. Murata K., Mitsuoka K., Hirai T., Walz T., Agre P., Heymann J.B., Engel A., and Fujiyoshi Y.// Nature. 2000. 407. 603.
10.
Хакимов
А.М., Рудакова М.А., Дорогиницкий М.М. и Филиппов
А.Ф.// Биофизика 2008. 53. 271.
11.
Pershin
S.// Phys. of Wave Phenomena. 2008. 16(1). 15.
12.
Bloembergen N., Parcell E., Pound V.// Phys.Rev.
1948. 78, 679.
13.
Лёше А. Ядерная индукция - М:, 1963г., 684 с. ( A.Losche,
Kerninduktion, Berlin: Verlag, 1961.).
14.
Farkas
A., Orthohydrogen, Parahydrogen,
and Heavy Hydrogen –
15.
Rothman L.S., Jacquemart
D., Barbe A., Benner D.Ch., Brick M., Brown L.R., Carleer M.R., Chackerian C.,
Chance Jr.K., Coudert L.H.,
Dana V., Devi V.M., Flaud J.-M., Gamache
R.R., Goldman A., Hartmann J.-M., Jucks K.W., Maki
A.G., Mandin J.-Y., Massie S.T., Orphal
J., Perrin A., Rinsland C.P., Smith M.A.H., Tennyson
J.,. Tolchenov R.N, Toth
R.A., Auwera J.V.,
16.
Limbach
H.-H., Buntkowsky G., Matthes
J., Grundemann S., Pery T.,
Walaszek B., and Chaudret
B. ChemPhysChem., 2006. 7. 551.
17. Buntkowsky G., Limbach H.-H., Walaszek B., Adamczyk A., Xu Y., Breitzke H., Schweitzer A., Gutmann T., Wachtler M., Frydel J., Emmler Th., Amadeu N., Tietze D. Mechanism of Ortho/Para-H2O Conversion in Ice // Z. Phys. Chem. 2008.222. 1049.
18. Бункин А.Ф., Нурматов А.А., Першин С.М. Когерентная четырехфотонная спектроскопия низкочастотных либраций молекул в жидкости // УФН 2006. 176. 883.
19. Бункин А.Ф., Нурматов А.А., Першин С.М., Хусаинова Р.С., Потехин С.А., Биофизика, послана в печать (2008).
20.
Bunkin
A.F., Pershin S.M., Nurmatov
A.A.// Laser Phys. Lett. 2006. 3(6), 275-277.
21.
Potekhin
22.
Pal S.K., Peon J., and Zewail Ahmed H.// PNAS, 2002. 99(17), 10964–10969.
23. Peon J., Pal S.K., and Zewail Ahmed H.// PNAS, 2002. 99(4), 1763–1768.
24. Zheng J.and Pollack Gerald H.// Phys. Rev. E 2003, 68, 031408.
25. Джеральд Поллак, Клетки, гели и двигатели жизни: новый, унифицирующий взгляд на клеточные функции, Редакторы: Бляхман Ф. А., Воейков Л. В., Сафронов А. П. – Екатеринбург: физический факультет, Уральский Гос. Университет им. А.М. Горького, Свердловская обл., 620083 Россия, Тел. +7 (343) 2613057. E-mail: felix.bljakhman@usu.ru
26.
Liou
Yih-Cherng, Tocilj Ante,
Davies Peter L. & Jia Zongchao.
Mimicry of ice structure by surface hydroxyls and water of a b-helix antifreeze
protein // Nature, 2000. 406 , 20.
27. Pershin S.M., Bunkin A.F., Lukyanchenko V.A., Nigmatullin R.R. “ Detection of the OH band fine structure in liquid water by means of new treatment procedure based on the statistics of the fractional moments” // Laser Physics Letters. 2007. 4(11), 809-813.
28.
Першин
С.М., Адикс Т.Г., Лукъянченко
В.А., Нигматуллин Р.Р., Потапов А.А., “Наблюдение спектральной компоненты льдоподобных
комплексов в ОН полосе воды при температуре от 13 до
29.
Pershin
S. M. Two Liquid Water // Physics of Wave Phenomena. 2005. 13(4),
192-208; Pershin S.M. Harmonic oscillations of the
concentration of H-bond in liquid water // Laser Physics 2006. V.16,
#7, p.1-7.
30.
Teixeira J., Bellissent-Funel M.C.,
Chen S.H. and Dorner B. // Phys. Rev. Lett. 1985. 54. 2681; Ruocco G. and Sette F. The
high-frequency dynamics of liquid water //J. Phys.: Condens.
Matter. 1999. 11. R259–R293.
31.
Jinesh
K.B., and Frenken J.W.M. Experimental Evidence for
Ice Formation at Room Temperature // Phys. Rev. Lett.
2008. 101. 036101.
32.
Bunkin
A.F., Pershin S.M. J. Raman Spectrosc.
33.
Kudryavtsev
A.B., Christopher G., Smith C.D., Mirov S.B., Rosenblum W.M., DeLucas L.J. The
effect of ordering of internal water in thaumatin and
lysozyme crystals as revealed by Raman method //
Journal of Crystal Growth 2000. 219. 102-114.
34.
Pershin
S. M., Bunkin A.F. Observation of temperature
evolution of relative concentration ortho/para
spin-isomers H2O by four-photon spectroscopy // Laser Physics,
35. Салихов К.М., 10 лекций по спиновой химии –Казань: УНИПРЕСС, 2000, 143с.
36. Зельдович Я.Б., Бучаченко А.Л., Франкевич Е.Л. Магнито-спиновые эффекты в химии и молекулярной физике //1988, УФН, 155(1), 3-44.
37. Michout X., Vasserot A.-M., Abouaf-Marguin L. Temperature and time effects on the rovibrational structure of fundamentals of H2O trapped in solid argon: hindered rotation and RTC satellite // Vibr. Spectrosc., 2004 34, 83-93.
38. Вебер С.Л., Багрянская Е.Г., Чаповский П.Л., О возможности обогащения ядерных спиновых изомеров молекул Н2О при помощи адсорбции // ЖЭТФ, 2006. 129(1), 86-95.
39. Звелто О., Принципы лазеров, под ред. Т.А. Шмаонова, СПб.: «Лань», 2008, 720с. Principles of Lasers, 4th ed. By Orazio Zvelto, Plenum Publishing Corp. (1998).
40. Захаров С.Д., Еремеев Б.В., Перов С.Н.// Кр. Сообщ. По физ. ФИАН, 1989. №1, 15.
41. Захаров С.Д., Иванов А.В., Светокислородный эффект в клетках и перспективы его применения в терапии опухолей // Квантовая электроника, 29(3), 192-214 (1999); S.D. Zakharov, A.V. Ivanov, Light-oxygen effect in the cells and the prospects for its application in tumour therapy.
42.
Захаров
С.Д., Иванов А.В., Вольф Е.Б., Данилов В.П., Мурина Т.М., Нгуен
К.Т., Новиков Е.Г., Панасенко Н.А., Перов С.Н., Скопинов С.А., Тимофеев Ю.П.// Структурные перестройки в
водной фазе клеточных суспензий и белковых растворов при светокислородном
эффекте, Квантовая электроника. 2003. 33(2), 149-162.
43.
Structural rearrangements in the
aqueous phase of cell suspensions and protein solutions induced by a
light-oxygen effect.// Quantum Electronics, 2003. 33(2), 149-162.
44.
Otsuka
I.and Ozeki S., Does
Magnetic Treatment of Water Change Its Properties?// Phys. Chemistry B. 2006.
110, 1509-1512.
45.
Ozeki
S.and Otsuka I. Transient Oxigen Clatrate-like Hydrate and
Water Networks Induced by Magnetic Fields.//Phys. Chemistry B, 2006. 110,
20067-20072.
46.
Bunkin
A. F., Pershin S. M. Four-wave
mixing spectroscopy of hydration layers in biopolymers and carbon nanotubes aqueous solutions // Journal of Raman
Spectroscopy, 2009.
47.
Титовец Э.П. Аквапорины человека и
животных. Фундаментальные и клинические аспекты.// Минск, «Белорусская наука»,