На эту статью можно ссылаться, ее адрес в интернете: www.biophys.ru/archive/congress2009/pro-p162.htm
О ВОЗМОЖНОЙ РОЛИ СПИНОВОЙ ПОДСИСТЕМЫ В МЕХАНИЗМЕ ДЕЙСТВИЯ ФАКТОРОВ НИЗКОЙ ИНТЕНСИВНОСТИ
А.В.Дроздов
198103, Россия, Санкт-Петербург, Институт аналитического приборостроения РАН,
Рижский пр-т д.26, E-mail: av@biophys.ru
Введение
Осознанный интерес к действию электромагнитных полей и неионизирующих электромагнитных излучений на биологические системы возник у человечества одновременно с осознанием факта их существования. Так, например, первые опыты по изучению воздействия электромагнитного излучения на людей были начаты Де Арсонвалем в 1891 году – через три года после открытия Герцем свободных электромагнитных волн и конструкции им первого излучателя. К настоящему времени эта область научных интересов уже сформировалась в одно из признанных направлений современного естествознания, называемого электромагнитобиологией. Основной целью электромагнитобиологических исследований является установление закономерностей и механизмов дистантного действия ЭМП на различные биологические системы.
И хотя развитие электромагнитобиологии привело к практическому применению ЭМП, вопрос «Какова роль естественных электромагнитных полей, существующих на Земле, в обеспечении функционирования различных организмов и их участии в эволюции?» до сих пор не имеет ответа. Между тем, к настоящему времени накоплено много свидетельств того, что некоторые электромагнитные поля природного происхождения (регистрируемые современными приборами на пределе возможности или даже не регистрируемые вовсе) представляют потенциальную угрозу для здоровья людей и стоят в одном ряду с такими климатическими факторами как температура, давление и т.п. Эти результаты свидетельствуют в пользу того, что изучение влияния ЭМП природного и антропогенного происхождения на жизненно важные физико-химические процессы является весьма актуальной задачей, значимость которой трудно переоценить.
В истории науки неоднократно констатировались явления или группы явлений, характеризующиеся тем, что очень слабые с привычной (энергетической) точки зрения, или даже вообще не поддающиеся регистрации воздействия вызывают весьма значительный по силе отклик биологических [1], а в некоторых случаях и абиогенных систем [2]. В роли таких воздействий могут выступать физические поля, электромагнитные или корпускулярные излучения очень низкой интенсивности, очень малые концентрации активных веществ, вносимых в систему [3], очень слабые акустические или силовые механические воздействия и т.д. В целом ряде случаев само воздействие вообще не определяется, регистрируются лишь статистически достоверные корреляции между внешне никак не связанными явлениями: например, теми или иными космическими событиями, с одной стороны, и статистикой заболеваемости или скоростью биохимических процессов - с другой [4, 5].
Однако, несмотря на результаты многолетних наблюдений и важность решаемых электромагнитобиологией задач, у значительной части научного сообщества до сих пор имеются сомнения в существовании самой проблемы. Отчасти, эти сомнения связанны с принципиальной невозможностью действия электромагнитных полей и излучений низкой интенсивности[1] на биосистемы. Такая невозможность вытекает из современных термодинамических подходов, используемых в современной биохимии. Часто биологические эксперименты по изучению действия низкоинтенсивных ЭМП состоят в наблюдении связи между физико-химическими характеристиками действующего фактора/факторов и вызываемого биологического отклика. В результате такого подхода выяснить механизмы действия слабых ЭМП на уровне отдельных биохимических реакций, являющихся основой жизнедеятельности любого организма, не представляется возможным. Поскольку в ходе планирование таких экспериментов нет возможности оценить степень вовлеченности различных уровней организации живой системы (биохимического, физиологического и т.п.), многие важные факторы остаются вне поля зрения исследователей. Все эти объективные трудности делают задачу предложения и обоснования общей концепции действия факторов низкой интенсивности на биологические системы весьма трудной.
При этом стоит особо подчеркнуть, что в рамках научного направления «Действие слабых и сверхслабых поле и излучений/факторов низкой интенсивности» принципиальным является поиск «низкоэнергетического» механизма. Механизма способного объяснить имеющиеся проблемы и экспериментальные результаты на современном научном уровне, при этом по возможности не нарушая правила «Лезвие Оккама»[2].
Представляется, что на сегодняшний день такая возможность существуют. Целью настоящей работы является попытка посмотреть на проблему действия факторов низкой интенсивности с квантово-механических позиций. Представляется весьма полезным и интересным проведение всестороннего анализа степени вовлеченности спиновых систем в формирование отклика на действие слабых и сверхслабых полей и излучений (далее факторов низкой интенсивности) как биологическими, так и небиологическими системами. Расширение круга рассматриваемых систем, возможно, позволит посмотреть на «биологическую проблему» с более общих позиций.
Часть I
Пытаясь разобраться в механизмах действия факторов низкой интенсивности, прежде всего, необходимо выбрать первоначальный уровень рассмотрения, масштаб с которого стоит начать изучение проблемы. Правильность выбора масштаба должна позволить выявить мишени действия этих факторов, что в последствие должно сильно помочь при формировании общей концепции.
При изучении действия внешних воздействий на биосистемы мы, как правило, оперируем такими понятиями как биоэффект, величина биоэффекта. В основе такого подхода лежит изучение разницы протекания каких-либо биологических процессов в экспериментальной группе, на которую производится низкоинтенсивное воздействие, с контрольной группой, на которую такого воздействия не происходит. Фактически, наши внешние воздействия направлены на изменение скорости биохимических реакций лежащих в основе наблюдаемых биоэффектов. Таким образом, представляется, что изучение действия низкоинтенсивных воздействий может быть обращено к рассмотрению химических реакций - биологически значимых для живых систем и структурно образующих в не живых.
Такой выбор означает, что в дальнейшем будет рассмотрено центральное событие в химии – образование химической связи. Стоит заметить, что само это химическое событие весьма непродолжительно (порядка 10-11 – 10-13 с). Оно составляют ничтожно малую часть от общего времени химической реакции; почти все время необходимо на подготовку события – встреча реагентов, организация их подходящей ориентации и энергетической накачки. Именно «подготовительная» стадия является причиной разнообразия временного масштаба химического процесса – от медленных до быстрых.
Выбор химической реакции в качестве основного события нашего внимания позволяет сформулировать задачи, решение которых поможет продвинуться в понимании механизмов действия факторов низкой интенсивности. К первоочередной задаче можно отнести вопрос: «Какие факторы управляют химической реакцией?». Ответ на этот вопрос позволяет определить направление дальнейших исследований. На сегодняшний день таких факторов два. Это энергия и угловой момент реагентов.
Хорошо известно, что химическими реакциями правит энергия – через энергию электронов, реакционную способность, активационные барьеры. Энергетический запрет означает, что система должна обладать энергией, не меньшей, чем высота потенциального барьера. Барьера, который разделяет два события «встреча реагентов» и «образования продукта реакции». Все принципы управления химическими реакциями исходят из энергетической догмы: управлять реакцией значит играть на ее энергоемкости – снимать энергетические барьеры, когда реакцию нужно стимулировать или, напротив, воздвигать их, когда реакцию нужно подавлять. К настоящему моменту на «энергетическом» поле химия достигла высокого уровня совершенства. Однако эта победа оказалась не окончательной. На арене появился и все более утверждается в своих правах, второй запрет – угловой.
Суть углового запрета заключается в том, что угловой момент в химических реакциях строго сохраняется. Этот фундаментальный закон означает, что химические реакции разрешены только для таких угловых состояний реагентов, угловой момент которых совпадает с угловым состоянием продукта, и строго запрещены, если требуется его изменение. Это относится ко всем угловым моментам – вращательному моменту молекулы, орбитальному электронному моменту, собственному моменту (спину) электрона и собственному ядерному моменту (ядерному спину). Запреты химических реакциями по угловому моменту менее очевидны, хотя их строгость значительно превосходит строгость энергетических запретов. Например, существуют реакции, в которых нет необходимости переваливаться через энергетический барьер, в этих реакциях возможно перемещение одного из реагентов под барьером, без энергетической накачки.
Именно обзор проявлений второго запрещающего фактора и предлагается положить в основу исследования механизмов действия факторов низкой интенсивности, т.е. рассмотреть взаимодействие угловых моментов (спинов) с внешними слабым воздействием, как в живой, так и не живой природе.
Для большей наглядности наших рассуждений обратимся к давно волнующему науку вопросу «Влияет ли магнитное поле на скорость химической реакции?». Ответ на этот вопрос давно интересует физиков, химиков и биологов — всех, кому приходится изучать влияние внешних факторов на превращения веществ. Особенно важен ответ на этот вопрос для биологов, так как биохимические процессы всегда происходили и происходят в магнитном поле Земли. И если слабое магнитное поле Земли способно воздействовать на химические реакции, то вполне возможно, что это же поле может оказывать существенное влияние и на жизнедеятельность земных организмов.
Если обратиться к истории вопроса, то можно найти много работ, авторам которых удавалось наблюдать действие внешнего магнитного поля на ход химических процессов; но очень неприятная особенность этих результатов заключалась в их плохой повторяемости. Кроме того, создавалась вообще парадоксальная ситуация: с одной стороны, накапливались все новые и новые факты, свидетельствующие о химической активности магнитного поля, но с другой стороны, росли и сомнения в принципиальной возможности подобного эффекта.
Простое накопление фактов, полученных в ходе экспериментов, поставленных наугад и не нацеленных на доказательство или опровержение какой-либо конкретной гипотезы, часто не дает определенных результатов. Дело в том, что те или иные явления могут наблюдаться лишь в строго определенных условиях, и если эти условия не соблюдены, то и результаты опытов будут отрицательными. Но когда механизм явления вообще неизвестен, то в одних опытах эффект может наблюдаться, а в других — нет. Все это приводит к убийственному для каждого экспериментатора результату – невоспроизводимости.
С другой стороны, рассматривая проблему действия магнитных полей с энергетических позиций понятно, что для того чтобы свойства частиц участвующих в реакции, изменились под действием внешнего поля, внесенная извне энергия должна быть соизмеримой с энергией теплового движения частиц или превосходить ее. Однако энергии магнитного взаимодействия и теплового движения становятся соизмеримыми лишь при использовании магнитных полей напряженностью Н=106 эрстед. Так, например магнитное поле напряженностью Н=1,4×104 эрстед изменяет энергию свободного радикала так же, как и изменение его температуры на 1 °С. Поэтому надежды на сколь бы то ни было заметное действие магнитного поля на молекулярном уровне до недавнего времени связывались только с реакциями при очень низких температурах или в сверхсильных магнитных полях. Для сравнения энергетических масштабов в таблице 1 приведены различные энергетические величины.
Таблица 1.
|
Энергия |
Единицы измерений (Дж/моль) |
|
Энергия диссоциации Н2О®Н+ОН |
4,9×105 |
|
Тепловая энергия кТ (Т=300 К) |
2400 |
|
Магнитная (зеемановская) энергия электрона в поле Н=3000 Гс |
3,4 |
|
Магнитная энергия электрона в поле протона (сверхтонкое взаимодействие в атоме водорода) |
0,56 |
В целом отношение к проблеме, действия магнитных полей, бытующих в науке и по сей день, хорошо отражает статья «Магнетохимия» в Краткой химической энциклопедии (1963 год): «...При не очень низких температурах энергии атомов и небольших молекул вещества непрерывно подвергаются столь сильным случайным флуктуациям вследствие хаотического теплового движения, что изменения энергии, вызываемые обычными (до 104 эрстед) магнитными полями, оказываются незаметными. Поэтому влияние такого рода магнитных полей на скорость химических реакций является незначительным по сравнению с влиянием температуры. Заметного эффекта можно было бы, по-видимому, ожидать для небольших молекул лишь при магнитных полях, в сотни и тысячи раз более сильных, чем обычные поля, создаваемые в лабораторных условиях. Однако подобного рода сверхсильные поля в настоящее время удается поддерживать лишь в течение очень коротких промежутков времени (~10-3—10-4 с), так что опытная проверка влияния магнитных полей на кинетику химических реакций до сих пор не производилась». Это с одной стороны.
Но с другой стороны, если предположить, что наблюдаемые экспериментальные результаты по воздействию магнитных полей на химические реакции, даже при не соответствии энергии магнитного поля энергетическим масштабам реакции, все же имеют место, то необходимо понять как такое, может быть. Предполагая саму возможность действия магнитного поля на химические системы, надо понимать следующее: такое действие может быть связано лишь с изменением энергии каких-то отдельных частиц вещества. Такими частицами способны служить либо сами молекулы, либо их части – свободные радикалы. В принципе каждый реагент в химической реакции является свободным радикалом, поскольку содержит, по меньшей мере, один неспаренный электрон, а электрон обладает моментом количества движения, или спином, и связанным с ним магнитным моментом. Теоретически, магнитное поле может внести в расположение магнитных частиц некоторый порядок, повлияв тем самым на скорость химического процесса, однако тепловое движение приводит к тому, что магнитные моменты свободных радикалов ориентируются по направлениям в пространстве как попало, или, как говорят, с одинаковой вероятностью. В этом как раз и есть проявление «проблемы кТ».
Попробуем подробнее разобраться с возможность действия магнитного поля на отдельный магнитный момент свободного радикала участвующего в наиважнейшем для нас событии – образование химической связи.
Часть II
В качестве примера возьмем реакцию образования двухатомной молекулы М. В этом случае, аналогом свободных радикалов будут являться атомы с неспаренными электронами. Схематически это можно изобразить так (на схеме кружками обозначены неспаренные электроны):
![]()
Вообще говоря, в дальнейшем нас будет интересовать действие магнитного поля не на отдельный свободный радикал, а на так называемую радикальную пару, т.е. пару образованную реагентами химической реакции. Схема реакции примет вид:
![]()
Процессы, обозначенные горизонтальными стрелками на схеме, связаны с молекулярным движением (диффузией) радикалов и химическими процессами - диссоциацией молекул и рекомбинацией пары. Промежуточное состояние, выделенное скобками, и есть радикальная пара (РП).
Согласно теории валентной связи Гайтлера-Лондона валентная связь создается двумя электронами. Поскольку электрон имеет спин S=1/2, то суммарный спин двух электронов может равняться нулю или единице (S=0, 1). Состояние с суммарным спином нуль (S=0) называется синглетным, в этом состоянии спины двух электронов ориентированы в противоположные стороны. Состояние с суммарным спином единица (S=1) называется триплетным, в этом состоянии спины двух электронов одинаково ориентированы. Суммарный спин двух электронов участвующих в образовании химической связи жестко связан с пространственным распределением электронов. Согласно принципу Паули, в одной точке пространства не могут находиться одновременно два электрона в одинаковом спиновом состоянии. Это означает, что в синглетном состоянии два валентных электрона могут одновременно находится в пространстве между двумя атомами, а в триплетном – это запрещено принципом Паули. Иными словами если сталкивающиеся радикалы находится в синглетном состоянии, тогда образование химической связи возможно. Такая реакция, как правило, протекает быстро, эффективно и безактивационно, т.е. энергия активации реакции близка к нулю. Если же сталкивающиеся радикалы находится в триплетном состоянии, то образование молекул - запрещено.
Исходя из выше сказанного, следует несколько уточнить модель реакции, введя в рассмотрение синглетные и триплетные состояния радикальной пары. В результате мы приходим к следующей схеме (вертикальными стрелками на этой схеме обозначена спиновая динамика в РП, синглет-триплетные переходы):
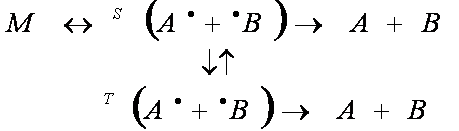
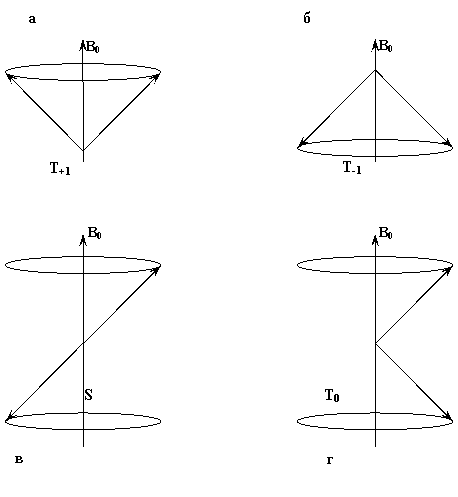
Перейдем к рассмотрению непосредственно самой реакции, но уже с позиций спиновой динамики. В качестве примера возьмем реакцию рекомбинации радикалов, которые зарождены по отдельности вдали друг от друга и реагируют при случайных столкновениях. В этом случае спиновое состояние пары сталкивающихся радикалов случайно. При случайной встрече двух радикалов в растворе их спины не коррелированны. К моменту встречи суммарный спин двух неспаренных электронов радикалов может быть 0 или 1, причем состояние с S=1, в интересующем нас случае наличия магнитного поля, трехкратно вырождено (возможны три проекции спина на направление внешнего магнитного поля S=+1, 0, -1), а состояние с S=0 невырождено (рис.1). В соответствии с этими степенями вырождения статистический вес встреч двух радикалов в синглетном состоянии равен 1/4, а соответственно статистический вес встреч в триплетном состоянии равен 3/4. В подавляющем большинстве случаев основное состояние продукта рекомбинации двух радикалов является сиглетным, и поэтому в соответствии с правилом Вигнера (сохранения спинового состояния реагентов в элементарном химическом акте) следует ожидать, что только 1/4 столкновений может привести к протеканию реакции рекомбинации, а 3/4 столкновений могут привести только к протеканию других конкурирующих реакций (реакциям с растворителем, с радикальными ловушками, изомеризации или диссоциации радикала и т.д.).
|
|
Рис.1. Схематическое представление возможных комбинации взаимного расположения спинов электронов в радикальной паре. На Рис.1(в) показаны радикалы, проекции магнитного момента которых оказались при встрече направленными в различные стороны, причем прецессия этих радикалов происходит в противофазе. В этом случае как суммарный спин системы двух радикалов, так и суммарная проекция их магнитных моментов оказывается равной нулю. Это означает, что радикальная пара находится в синглетном состоянии, из которого возможна реакция рекомбинации. На Рис.1(а,б,г) суммарный спин пары равен единице. На Рис.1(г) проекции магнитных моментов радикалов направлены в различные стороны и суммарная проекция на направление магнитного поля (продольная поляризация) равна нулю. Однако прецессия спинов происходит синфазно, и, таким образом, суммарный спин поперек направления магнитного поля (поперечная поляризация) равен 1. На Рис.1(а), проекции магнитных моментов обоих радикалов направлены по направлению внешнего магнитного поля, а на Рис.1(б) - против внешнего поля. Таким образом, при случайной встрече радикалов реализуется один из четырех вариантов спинового состояния радикальной пары - синглетное состояние S (s=0, ms=0) и три триплетных состояния T+, T0, T-(s=1, ms = -1,0,1). |
Из приведенного выше примера следует, что спиновое состояние реагирующих частиц в значительной степени определяет направление реакции и состав образующихся продуктов. Такое рассмотрение было бы исчерпывающим, если бы спиновое состояние реагирующих частиц не могло изменяться. Для понимания вероятности протекания реакции по тому или иному направлению необходимо рассмотреть возможность изменения спинового состояния радикальной пары.
Магнитное поле вносит в расположение частиц определенный порядок, а тепловое движение приводит к тому, что магнитные моменты свободных радикалов ориентируются по направлениям в пространстве как попало, или, как говорят, с одинаковой вероятностью. Поэтому пытаясь разобраться в спиновой динамике, мы должны, прежде всего, понять каково соотношение между организующими факторами и дезорганизующими.
Скорость изменения спинового состояния молекул и атомов принято характеризовать величинами T1 (время продольной или спин-решеточной релаксации) и T2 (время поперечной или спин-спиновой релаксации). Проиллюстрируем эти характеристики на примере. Пусть в образце содержится ансамбль частиц со спином 1/2. Они могут быть ориентированы либо по, либо против поля. В магнитном поле энергия этих состояний различна. Спины в образце заселяют эти два состояния в соответствии с распределением Максвелла-Больцмана. При этом разность равновесных максвелл-больцмановских заселенностей двух состояний характеризует равновесную намагниченность образца. Предположим, что с помощью внешнего воздействия часть спинов переходит в другое состояние и, таким образом, изменяется намагниченность образца. Нарушенное распределение начнет релаксировать к равновесию. Как долго могут существовать такие поляризованные системы, не переходя в равновесное состояние (в котором концентрации синглетных и триплетных удовлетворяет соотношению 1:3)? Это время (в органических и значительной части неорганических радикалах эти времена лежат в интервале 10-5 - 10-8 с) как раз и определяется временами Т1 и T2, т.е. скоростью с которой энергия теплового хаотического движения окружающих частиц приводит спиновую подсистему к равновесному распределению.
Если продолжительность жизни пары радикалов t меньше Т1 и T2 (условие t£T1,T2 во многих случаях действительно выполняется, так как в типичном случае t ~10-9 с), то тепловое движение просто не успеет оказать на частицы никакого влияния — их магнитные моменты либо сохранят свою взаимную ориентацию, либо изменят ее по причинам, не связанным с хаотическим движением других молекулами. Таким образом, магнитные моменты электронов на протяжении всего времени существования радикальной пары не имеют связи с тепловым движением и ведут себя как бы при абсолютном нуле температуры. Так устраняется главный конкурент «слабого» внешнего магнитного поля — тепловое движение, и тем самым решается пресловутая «проблема кТ».
Стоит заметить, что очень важно иметь оптимальное время жизни РП. В невязких растворах время жизни РП довольно короткое, меньше или порядка наносекунд, и поэтому для реакций в невязких жидкостях магнитные эффекты слабо выражены. Но очень большое время жизни РП тоже нежелательно, так как при этом независимо от индукции магнитного поля успеет установиться статистическое распределение РП по синглетному и триплетному состояниям. Оптимальным является время жизни РП, которое соизмеримо со временем синглет-триплетных переходов в РП. В обычных условиях это время невелико, однако изменяя условия эксперимента (вязкость и температуру), им можно управлять. Например, для реакции РП в мицеллах реализуется такое оптимальное время жизни. Таким образом, в жидком растворителе у радикальной пары есть время для спиновой эволюции, т.е. для перехода из одного спинового состояния в другое.
Все эти рассуждения имеют одну единственную цель - подвести к пониманию того, что найдя способ/способы увеличения вероятность перехода (спиновой конверсии) свободных радикалов в синглетное состояние, мы неминуемо придем к изменению скорости химической реакции, а значит и к – биологическому эффекту. К сожалению, объем данной работы не позволяет подробно остановиться на причинах, приводящих к спиновой динамике. Скажем лишь, что у достаточно молодой науки «Спиновая химия» есть весьма серьезные достижения [6]. Уже сегодня ей известны механизмы позволяющих осуществлять спиновую конверсию меняя тем самым масштаб химической реакции. К ним относятся: Dg-механизм, СТВ-механизм, СВЧ стимуляция. Получены результаты, которые свидетельствуют о том, что:
· «…Ключевая биохимическая реакция – синтез АТФ – является спин-селективной; она происходит в спин-селективном нанореакторе, который может функционировать как молекулярный приемник резонансных электромагнитных волн. Изменяя спин ион-радикальной пары, они переключают реакцию между спин разрешенным и спин запрещенным каналами, регулируя выход АТФ…» [7];
· «…скорость спиновой релаксации в слабых полях может на порядки отличаться от скорости релаксации в сильных полях. Причем эту особенность нельзя объяснить в рамках сильнопольной теории» [8];
· «…скорость спиновой релаксации, обусловленной спин-вращательным взаимодействием, увеличивается в слабых полях. …в сильных магнитных полях (В0) ЭПР-переходы разрешены только при В1^В0, в то время как в слабых полях разрешены также переходы при В1||В0» [8].
Таким образом, на вопрос о возможности влияние магнитного поля на скорость химической реакции спиновая химия дала утвердительный ответ. Причем не просто сказала «Да, влияет!», а экспериментально показала возможность действия магнитного поля на биологически значимую реакцию - синтез АТФ.
С биологических позиций из этого результата может следовать, что внешнее магнитное поле Земли синхронизирует работу живых клеток любого биологического организма. Действие магнитного поля, проникающего непосредственно на уровень каждой клетки, направлено на магнитный момент неспаренного электрона свободного радикала (ядерный магнитный момент, орбитальный магнитный момент и т.п.). Свободные радикалы становятся своего рода «антеннами» внутри нас, которые могут воспринимать управляющее воздействие от внешних факторов. Управляя спиновыми состояниями реагирующих частиц можно изменять временной масштаб реакции – управлять скоростью химического взаимодействия, изменяя тем самым степень биоэффекта.
Поскольку энергия «переключения» между спиновыми состояниями реагирующих частиц в магнитном поле Земли очень мала (на несколько порядков ниже энергии тепловых колебаний), а временные масштабы процессов на спиновом уровне короче времени тепловой релаксации, то нетрудно себе представить низкоэнергетический механизм действия факторов низкой интенсивности без привлечения энергоинформационных представлений.
И в конце еще одно замечание. В нашем рассмотрении в фокусе внимания была рекомбинация радикалов, поэтому речь все время шла о радикальной паре. Но все наши рассуждения применимы к любым реакциям описанного выше типа. В качестве А и В могут выступать любые частицы. Например, это могут быть атомы, электрон-дырочная пара в полупроводниках, пара радикал + триплетная молекула (скажем, R + О2) и т.д. и т.п.
Часть III
Каждый исследователь, занимающийся изучением механизмов действия на биосистемы слабых воздействий, в конечном итоге приходит к мысли о том, что вода, являющаяся основой всего живого, должна играть важную роль в этих процессах. Попробуем посмотреть и на воду с квантово-механических позиций.
В классической механике одинаковые частицы (например протоны), несмотря на тождественность их физических свойств, не теряют своей «индивидуальности». Действительно, если следить за движением частиц по каждой из своих траекторий, то в любой момент времени частицы можно идентифицировать.
В квантовой же механике положение совершенно меняется. В силу принципа неопределенности понятие траектории полностью теряет смысл. Можно сказать, что в квантовой механике одинаковые частицы полностью теряют свою «индивидуальность». Одинаковость частиц по их физическим свойствам приводит к их полной неразличимости. Принцип неразличимости одинаковых частиц играет одну из основных ролей при квантово-механическом исследовании систем, состоящих из одинаковых частиц. В связи с этим принципом, для системы, состоящей из двух и более одинаковых частиц, имеется всего две возможности: волновая функция всей системы либо симметрична, либо антисимметрична.
Применение принципа неразличимости к молекуле H2O приводит к тому, что молекулы воды должны быть разделены на два вида отличающихся друг от друга ориентировкой спинов протонов и ротационными квантовыми числами всей молекулы. Молекулы воды, в которых ядерные магниты антипараллельны (¯) называются пара-H2O. Молекулы при этом могут иметь только четные ротационные квантовые числа (0, 2, 4…). У орто-H2O ядерные магниты параллельны () и ротационные квантовые числа могут принимать только нечетные значения (1, 3, 5…).
В то время как несимметрично построенная молекула, например HCl, свободно может из состояния вращения с четным квантовым числом перейти к вращению с нечетным, у молекулы, построенной из одинаковых ядер, как например H2O, подобный переход спонтанно не происходит. Взаимная связь между ориентировкой ядерных спинов и ротационными квантовыми числами, а также запрещение перехода орто↔пара (спиновая конверсия) вытекают непосредственно из квантовой механики и хорошо подтверждаются спектроскопическими экспериментальными результатами.
Различие между пара-H2O и орто-H2O не сводится только к обозначению определенной внутренней структуры, а в результате запрета перехода пара-H2O ↔ орто-H2O указывает на существование двух, во многих отношениях отличных и практически не комбинирующие друг с другом, типов молекул.
Одним из основных различий между пара- и орто- спин-изомерами воды является то, что у первой есть состояние, в котором молекула не вращается (ротационное квантовое число равно нулю). В то время как у орто- - такого состояния нет, она всегда «крутится» подобно юле. Это различие приводит к тому, что у разных спиновых изомеров воды должна быть разная степень адсорбции на поверхности. Пара-H2O может участвовать в образование структур посредством водородных связей (в частности льдоподобные структуры), в то время как у орто-H2O образование связей возможно только за счет электростатических сил – ван-дер-ваальсовых взаимодействий. Так например, давно обсуждаемое биологами [9] отличие внутриклеточной (связанной) воды от обычного водно-солевого раствора, может быть объяснено с позиций пара-H2O и орто-H2O.
С другой стороны, хорошо известно, что биологическая активность биоструктур во многом определяется степенью их гидратации. Можно предположить, что изменение соотношения орто- /пара- воды, входящей в биологическую систему, должно приводить к изменению степени гидратации биоструктур, а это, в свою очередь, - к изменению их биологической активности. Таким образом, стимуляция процессы спиновой конверсии пара-H2O ↔ орто-H2O должна приводить к изменению масштабов биологических процессов, ускоряя или замедляя их.
Говоря о различной степень адсорбции пара-H2O и орто-H2O нельзя не вспомнить о получивших в последнее время широкое развитие солитонных представлениях [10, 11]. В этих теориях рассматриваются коллективные состояния квантовых подсистем и решаются уравнения движения энергии в биологических системах. В этой связи хотелось бы сказать о том, что в теории солитонов степень связи молекул, вовлеченных в коллективное взаимодействие с поверхностью биомембран и друг с другом должно играть наиважнейшую роль.
Заключение
Приведенные выше рассуждения указывают на то, что при поиске механизма, отвечающего за отклик различных систем (как живых, так и не живых) на действие факторов низкой интенсивности, спиновые подсистемы - играть важну, а может и ключевую роль. Рассмотрение проблемы «слабые и сверхслабые поля и излучения» с квантово-механических позиций позволяет:
· решить «проблему kT»;
· предложить низкоэнергитечиский механизм действия факторов низкой интенсивности;
· отказаться от понятия «энерго-информационное воздействие».
Похоже, маленький спин – играет БОЛЬШУЮ роль!!!
Литература.
1. Труды международного конгресса «Слабые и сверхслабые поля и излучения в биологии и медицине» //СПб 1997, 2000, 2003. 2006.
2. С.Э.Шноль, В.А.Коломбер, Э.В.Пожарский, Т.А.Зенченко, И.М.Зверева, А.А.Конрадов, О реализации дискретных состояний в ходе флуктуаций в макроскопических процессах. // УФН, 1998, т.168, № 10, с.1129 - 1140.
3. Е.Б.Бурлакова, Особенности действия сверхмалых доз биологически активных веществ и физических факторов низкой интенсивности // Рос. хим. журн., 1999. Вып. 5. С. 3-11.
4. А.Л.Чижевский, Земное эхо солнечных бурь // М, Мысль, 1978.
5. Б.М.Владимирский, Н.А.Темурьянц Влияние солнечной активности на биосферу-ноосферу (Гелиобиология от А.Л.Чижевского до наших дней). // М.: Изд-во МНЭПУ, 2000. 374 с.
6. Бучаченко А.Л., Молин Ю.Н., Сагдеев Р.З., Салихов К.М. Магнитные и спиновые эффекты в химических реакциях. – М: Наука, 1984.
7. А.Л.Бучаченко, Новая изотопия в химии и биохимии. // М., Наука, 2007.
8. М.В.Федин, С.Р.Шакиров, П.А.Пуртов, Е.Г.Багрянская, Электронная спиновая релаксация радикалов в слабых магнитных полях. // Известия Акад.наук сер.хим, 2006, №10, стр.1643-1655.
9.
G.H.Pollack:
Cells, Gels and the Engines of Life: A New, Unifying Approach to Cell Function.
// Ebner & Sons,
2001.
10. А.С.Давыдов. Солитоны в биологических системах. // Киев, Наукова думка, 1988, 364с.
11.
E. Del Giudice, S.Doglia, M.Milani, G.Vitiello. A quantum
field theoretical approach to the collective behavior of biological system. // Nuclear Physics 1985, B251, [FS 13], p.375-400.
About possible role
of spin status in mechanism effected of low power activity factors
A.V.Drozdov
Institute for Analytical Instrumentation of the
Saint-Petersburg, Rizhsky
Pr. 26, E-mail: av@biophys.ru
[1] Под низкоинтенсивными воздействиями понимаются такие воздействия, энергия которых много меньше характерных энергий биохимических реакций. Часто в литературе этот энергетический дисбаланс внешнего воздействия и термодинамического равновесия в биологических системах называется «проблемой kT».
[2] «Бритва (лезвие) О́ккама» — методологический принцип. В упрощенном виде он гласит: «Не следует привлекать новые сущности без самой крайней на то необходимости».